Research Article
Naeem Abdul Ghafoor ![]() 1 , Beata Sitkowska
1 , Beata Sitkowska ![]() 2
2
1Department of Molecular Biology and Genetics, Faculty of Science, Muğla Sıtkı Koçman University, 48000 Muğla, Turkey
2Department of Animal Biotechnology and Genetics, Faculty of Animal Breeding and Biology, Bydgoszcz University of Science and Technology, 85-084 Bydgoszcz, Poland.
Abstract. Mastitis is the single most expensive disease among cattle in the dairy industry and environmental pathogens such as Staphylococcus aureus and Staphylococcus warneri are among the most common culprits. Recent studies had shed the light on how such pathogens utilize lipolysis mechanisms to evade their host's immune response. In this study, computational drug discovery approaches were deployed to investigate human FDA-approved drugs that hold the potential of serving as inhibitors of lipase activity in the former 2 pathogens. Comprehensive computational analysis involving molecular docking, nanoscale molecular dynamics, and in silico binding free energy estimation has shown that Drospirenone, a unique progestogen with anti-mineralocorticoid properties commonly used in human birth control pills holds potential inhibitory activity against the lipase of Staphylococcus warneri as it had shown to form several stable hydrophobic interactions and hydrogen bonds with the formers lipWY lipase enzyme.
Keywords: mastitis, cattle milk, Drospirenone, drug repurposing, molecular dynamics
The global dairy industry was estimated to worth around 720 billion USD in 2019 and is expected to be worthed over a trillion USD by 2024 [Shahbandeh 2021Shahbandeh, M. (2021). Global dairy industry – statistics & facts. Statista. https://www.statista.com/topics/4649/dairy-industry/ Google Scholar]. With such high evaluation, the risks involved in the industry also increases, factors such as the milk yield, its quality, and composition especially affect the industry’s profitability with high margins. Somatic Cell Count (SCC) which mainly consists of leukocytes is one of the critical parameters used to assess the quality of milk as it is directly correlated to the health of the cattle’s udder, hence, higher SCC implies a higher degree of udder inflammation which reflects a lower milk quality [Cinar et al. 2015Cinar, M., Serbester, U., Ceyhan, A., Gorgulu, M. (2015). Effect of Somatic Cell Count on Milk Yield and Composition of First and Second Lactation Dairy Cows. Ital. J. Anim. Sci., 14(1), 3646. https://doi.org/10.4081/ijas.2015.3646]. SCC has also been used as an early indicator for the diagnosis of sub-clinical and clinical mastitis in cattle, cattle with less than 200,000 SCC · cm–3 are considered healthy whereas any increase above this range indicates an increased risk for mastitis [El-Tahawy and El-Far 2010El-Tahawy, A. S., El-Far, A.H. (2010). Influences of somatic cell count on milk composition and dairy farm profitability. Int. J. Dairy Technol., 63(3), 463–469. https://doi.org/10.1111/j.1471-0307.2010.00597.x]. Mastitis is also the single most expensive disease within the dairy industry, resulting in a loss of around 6% of the total production value annually within the industry due to several factors such as diagnosis, treatment, recovery, and yield loss [Gill et al. 1990Gill, R., Howard, W.H., Leslie, K.E., Lissemore, K. (1990). Economics of Mastitis Control. J. Dairy Sci., 73(11), 3340–3348. https://doi.org/10.3168/jds.S0022-0302(90)79029-7, Miller et al. 1993Miller, R.H., Paape, M.J., Fulton, L.A., Schutz, M.M. (1993). The Relationship of Milk Somatic Cell Count to Milk Yields for Holstein Heifers After First Calving. J. Dairy Sci., 76(3), 728–733. https://doi.org/10.3168/jds.S0022-0302(93)77396-8, Shim et al. 2004Shim, E.H., Shanks, R.D., Morin, D.E. (2004). Milk Loss and Treatment Costs Associated with Two Treatment Protocols for Clinical Mastitis in Dairy Cows. J. Dairy Sci., 87(8), 2702–2708. https://doi.org/10.3168/jds.S0022-0302(04)73397-4].
Besides posing the risk of sub-clinical and clinical mastitis, higher levels of SCC are also associated with abnormal changes in milk’s protein, fat, and lactose compositions resulting in overall low-quality milk [Lindmark-Månsson et al. 2006Lindmark-Månsson, H., Bränning, C., Aldén, G., Paulsson, M. (2006). Relationship between somatic cell count, individual leukocyte populations and milk components in bovine udder quarter milk. Int. Dairy J., 16(7), 717–727. https://doi.org/10.1016/j.idairyj.2005.07.003]. Mastitis can be caused by environmental and genetic factors, however, over 90% of the cases are known to be caused by microbial agents (mainly bacterial) like Streptococcus spp., Enterobacteriaceae, or Staphylococcus spp. either directly or through feed, eventually causing pathological lesions and inflammation of the mammary glands [Bianchi et al. 2019Bianchi, R.M., Schwertz, C.I., de Cecco, B.S., Panziera, W., De Lorenzo, C., Heck, L.C., Snel, G.G.M., Lopes, B.C., da Silva, F.S., Pavarini, S P., Driemeier, D. (2019). Pathological and microbiological characterization of mastitis in dairy cows. Trop. Anim. Health Prod., 51(7), 2057–2066. https://doi.org/10.1007/s11250-019-01907-0, Puppel et al. 2020Puppel, K., Kalińska, A., Kot, M., Slósarz, J., Kunowska-Slósarz, M., Grodkowski, G., Kuczyńska, B., Solarczyk, P., Przysucha, T., Gołębiewski, M. (2020). The Effect of Staphylococcus spp., Streptococcus spp. and Enterobacteriaceae on the Development of Whey Protein Levels and Oxidative Stress Markers in Cows with Diagnosed Mastitis. Anim. an Open Access J. from MDPI, 10(9), 1591. https://doi.org/10.3390/ani10091591]. The immune system in cattle’s are more often well suited to defend against most of these pathogens, for example, the lysosome enzyme found in the cattle’s milk can also digest the peptidoglycan layer found in most bacteria’s, and lactoferrin (a glycoprotein), found in milk and other secretions of the cattle, can also kill some bacteria’s by hindering their iron intake pathways, however, pathogens as well develop sophisticated mechanisms to combat such defense responses, recent studies showed that lipase enzymes secreted by Staphylococcus aureus play a major role in the evasion of the immune system in cattle by hydrolyzing the bacterial-derived lipoproteins thereby promoting their survival in the cattle’s mammary glands [Pyörälä 2002Pyörälä, S. (2002). New Strategies to Prevent Mastitis. Reprod. Domest. Anim., 37(4), 211–216. https://doi.org/10.1046/j.1439-0531.2002.00378.x, Chen and Alonzo 2019Chen, X., Alonzo, F. (2019). Bacterial lipolysis of immune-activating ligands promotes evasion of innate defenses. Proc. Natl. Acad. Sci., 116(9), 3764–3773. https://doi.org/10.1073/pnas.1817248116]. The most common coping mechanism used against such pathogens is the use of antibiotics and probiotics, however, the use of such methods results in the development of resistant microbial agents overtime at best and results in bioconcentration of such residues in the cattle’s milk [Koba et al. 2018Koba, I.S., Lysenko, A.A., Koshchaev, A.G., Shantyz, A.K., Donnik, I.M., Dorozhkin, V.I., Shabunin, S.V. (2018). Prevention of Mastitis in Dairy Cows on Industrial Farms. J. Pharm. Sci. Res., 10(10), 2582–2585. Google Scholar].
Computational drug discovery has gained huge momentum in recent years, especially with the availability of supercomputers and the integration of graphic cards to accelerate computation processes which made many bioinformatics and computational research highly spontaneous and less time-intensive, virtual screening for identification of lead compounds, and molecular dynamics which is exceedingly computationally expensive but holds a great promise in providing atom-level insights for biomolecular interactions under synthetic conditions mimicking the physiological cellular environment, the progress of such models for simulating complex molecular systems has also lead to the 2013 Nobel prize in chemistry [Liu et al. 2007Liu, W., Schmidt, B., Voss, G., Müller-Wittig, W. (2007). Molecular Dynamics Simulations on Commodity GPUs with CUDA BT – High Performance Computing – HiPC 2007 (S. Aluru, M. Parashar, R. Badrinath, V.K. Prasanna (eds.); pp. 185-196). Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-540-77220-0\_20, Thiel and Hummer 2013Thiel, W., Hummer, G. (2013). Methods for computational chemistry. Nature, 504(7478), 96–97. https://doi.org/10.1038/504096a, Nobile et al. 2017Nobile, M.S., Cazzaniga, P., Tangherloni, A., Besozzi, D. (2017). Graphics processing units in bioinformatics, computational biology and systems biology. Brief. Bioinform., 18(5), 870–885. https://doi.org/10.1093/bib/bbw058]. Drug repurposing on the other hand exploits the same mechanisms of de novo drug discovery with the additional advantage of minimizing the computational time and expensiveness of new drug research by repurposing already FDA-approved drugs (drugs approved by the U.S. Food and Drug Administration for use in humans) against new targets, given that such drugs already passed most of the in vitro, in vivo and clinical trials, the risk of side effects and toxicity is limited and the only risk to asses is those of efficacy and distribution [Oprea et al. 2011Oprea, T.I., Bauman, J.E., Bologa, C.G., Buranda, T., Chigaev, A., Edwards, B.S., Jarvik, J.W., Gresham, H.D., Haynes, M.K., Hjelle, B., Hromas, R., Hudson, L., Mackenzie, D.A., Muller, C.Y., Reed, J.C., Simons, P.C., Smagley, Y., Strouse, J., Surviladze, Z., Sklar, L.A. (2011). Drug repurposing from an academic perspective. Drug Discov. Today Ther. Strateg., 8(3), 61–69. https://doi.org/10.1016/j.ddstr.2011.10.002, Pushpakom et al. 2019Pushpakom, S., Iorio, F., Eyers, P.A., Escott, K.J., Hopper, S., Wells, A., Doig, A., Guilliams, T., Latimer, J., McNamee, C., Norris, A., Sanseau, P., Cavalla, D., Pirmohamed, M. (2019). Drug repurposing: progress, challenges and recommendations. Nat. Rev. Drug Discov., 18(1), 41–58. https://doi.org/10.1038/nrd.2018.168]. Such approaches have been extensively exploited in the recent COVID-19 pandemics to accelerate the discovery of novel therapeutics against several targets of the SARS-CoV-2 [Bakowski et al. 2021Bakowski, M.A., Beutler, N., Wolff, K.C., Kirkpatrick, M.G., Chen, E., Nguyen, T.H., Riva, L., Shaabani, N., Parren, M., Ricketts, J., Gupta, A.K., Pan, K., Kuo, P., Fuller, M., Garcia, E., Teijaro, J.R., Yang, L., Sahoo, D., Chi, V., Huang, E., Vargas, N., Roberts, A.J., Das, S., Ghosh, P., Woods, A.K., Joseph, S.B., Hull, M.V., Schultz, P.G., Burton, D.R., Chatterjee, A.K., McNamara, C.W., Rogers, T.F. (2021). Drug repurposing screens identify chemical entities for the development of COVID-19 interventions. Nat. Commun., 12(1), 3309. https://doi.org/10.1038/s41467-021-23328-0].
In this study, computational drug repurposing methodologies such as molecular docking based virtual screening and molecular dynamics were applied to discover lead drugs among FDA-approved drugs that possess potential anti-microbial activity on two of the most common pathogens (Staphylococcus aureus and S. warneri) that are responsible for elevated SCC and mastitis risk in cattle.
The sequence for the lip1 gene (UniProt: Q6GDD3) of Staphylococcus aureus and the lipWY gene (UniProt: Q5DWE2) of Staphylococcus warneri was retrieved from the UniProt database [UniProt Consortium 2017UniProt Consortium (2017). UniProt: the universal protein knowledgebase. Nucleic Acids Res., 45(D1), D158-D169. https://doi.org/10.1093/nar/gkw1099]. The SWISS-MODEL tool was used to generate the 3D structure of each of the lipase enzymes via the homology modeling approach [Waterhouse et al. 2018Waterhouse, A., Bertoni, M., Bienert, S., Studer, G., Tauriello, G., Gumienny, R., Heer, F.T., de Beer, T.A.P., Rempfer, C., Bordoli, L., Lepore, R., Schwede, T. (2018). SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res., 46(W1), W296-W303. https://doi.org/10.1093/nar/gky427].
A dataset compromising of the structure of 1993 FDA-approved drugs were retrieved from the eDrug3D database [Pihan et al. 2012Pihan, E., Colliandre, L., Guichou, J.-F., Douguet, D. (2012). e-Drug3D: 3D structure collections dedicated to drug repurposing and fragment-based drug design. Bioinformatics, 28(11), 1540–1541. https://doi.org/10.1093/bioinformatics/bts186, Douguet 2018Douguet, D. (2018). Data Sets Representative of the Structures and Experimental Properties of FDA-Approved Drugs. ACS Med. Chem. Lett., 9(3), 204–209. https://doi.org/10.1021/acsmedchemlett.7b00462]. Each of the 1993 FDA-approved drugs was docked against each of the 2 lipases’ via QuickVina-W, a blind-docking optimized version of the original AutoDock Vina (v1.1.2) tool [Trott and Olson 2010Trott, O., Olson, A. J. (2010). AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem., 31(2), 455–461. https://doi.org/10.1002/jcc.21334, Hassan et al. 2017Hassan, N.M., Alhossary, A.A., Mu, Y., Kwoh, C.-K. (2017). Protein-Ligand Blind Docking Using QuickVina-W With Inter-Process Spatio-Temporal Integration. Sci. Rep., 7(1), 15451. https://doi.org/10.1038/s41598-017-15571-7]. The docking trials were performed using a grid box around the entire enzyme structure (blind docking protocol) with an exhaustiveness of 256 (exhaustiveness is the measurement of how many times a trial is reproduced to be selected, the higher its value, the more accurate the results and the more computationally expensive is the calculation). To reduce methodological bias, the docking trial for the top 5 hits for each lipase was repeated again with the same configurations and the top 3 docked poses for each lipase-lead (2 · 3 = 6 total simulation system) with the highest binding affinity was selected (3 drugs for each lipase enzyme, total of 6 drug candidates). The results for each docking trail are provided as a separate plain text file in supplementary data 1 and 2 (S1 and S2).
The molecular dynamics simulations for each of the 9 lipase-lead complexes were performed via the Nanoscale Molecular Dynamics (NAMD v2.14 CUDA) tool [Phillips et al. 2020Phillips, J.C., Hardy, D.J., Maia, J.D.C., Stone, J.E., Ribeiro, J.V., Bernardi, R.C., Buch, R., Fiorin, G., Hénin, J., Jiang, W., McGreevy, R., Melo, M.C.R., Radak, B.K., Skeel, R.D., Singharoy, A., Wang, Y., Roux, B., Aksimentiev, A., Luthey-Schulten, Z., Tajkhorshid, E. (2020). Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys., 153(4), 044130. https://doi.org/10.1063/5.0014475]. CHARMM36m force field was used for both the lipase enzymes and their respective leads, the topology and parameter files were prepared via CHARMM-GUI [Jo et al. 2008Jo, S., Kim, T., Iyer, V.G., Im, W. (2008). CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem., 29(11), 1859–1865. https://doi.org/10.1002/jcc.20945, Huang et al. 2017Huang, J., Rauscher, S., Nawrocki, G., Ran, T., Feig, M., de Groot, B.L., Grubmüller, H., MacKerell Jr, A.D. (2017). CHARMM36m: an improved force field for folded and intrinsically disordered proteins. Nat. Methods, 14(1), 71–73. https://doi.org/10.1038/nmeth.4067]. Each pair of the lipase-lead complex was immersed in a square box with explicit TIP3P water such that a distance of 5 Å was maintained between the edge of the box and the lipase-lead complex along each axis, the system was neutralized with Na+ and Cl– ions and their final concentration was maintained at 0.15 mol · L–1 which reflects the cellular ion concentration [Terry et al. 2011Terry, C.A., Fernández, M-J., Gude, L., Lorente, A., Grant, K.B. (2011). Physiologically Relevant Concentrations of NaCl and KCl Increase DNA Photocleavage by an N-Substituted 9-Aminomethylanthracene Dye. Biochemistry 50(47), 10375–10389. https://doi.org/10.1021/bi200972c]. Each system was minimized and equilibrated for 10 ns at 310.15 K (physiological temperature) with periodic boundary conditions, Langevin dynamics, particle mesh Ewald (PME) under the NVT ensemble. The equilibration was followed by 50 ns of production simulation at the same temperature under the NPT ensemble. The trajectory from the production simulation was utilized to calculate the root mean square deviation (RMSD) of the lipase backbone (Cα) from the beginning of the simulation (t = 0 ns) to its end (t = 50 ns), and root mean square fluctuation (RMSF) of the lipase enzymes against their corresponding ligands to assess their motion throughout the simulation. The configuration file for the equilibration and production simulations is provided in S3 (for peer reproducibility).
The lead drugs that formed stable complexes with their respective lipase enzymes were submitted to the TU Dresden’s protein-ligand interaction profiler (PLIP) webserver to predict their interactions, interactions such as hydrogen bonds, hydrophobic interactions, pi-stacking, etc were visualized in PyMol (open-source v2.5.0) and corresponding snapshots were provided [Salentin et al. 2015Salentin, S., Schreiber, S., Haupt, V.J., Adasme, M.F., Schroeder, M. (2015). PLIP: fully automated protein-ligand interaction profiler. Nucleic Acids Res., 43(W1), W443-W447. https://doi.org/10.1093/nar/gkv315, Schrödinger 2015Schrödinger, L. (2015). The PyMOL Molecular Graphics System, Version 1.8.. Google Scholar].
The binding energy of the lead drugs that formed stable complexes with their respective enzymes at 310.15 K was calculated via the PROtein binDIng enerGY prediction (PRODIGY) algorithm available at the Utrecht biomolecular interaction web portal, the estimated binding free energy was calculated as the average between the binding energy calculated from the molecular docking and the one predicted by PRODIGY algorithm [Xue et al. 2016Xue, L.C., Rodrigues, J.P., Kastritis, P.L., Bonvin, A.M., Vangone, A. (2016). PRODIGY: a web server for predicting the binding affinity of protein-protein complexes. Bioinformatics, 32(23), 3676–3678. https://doi.org/10.1093/bioinformatics/btw514].
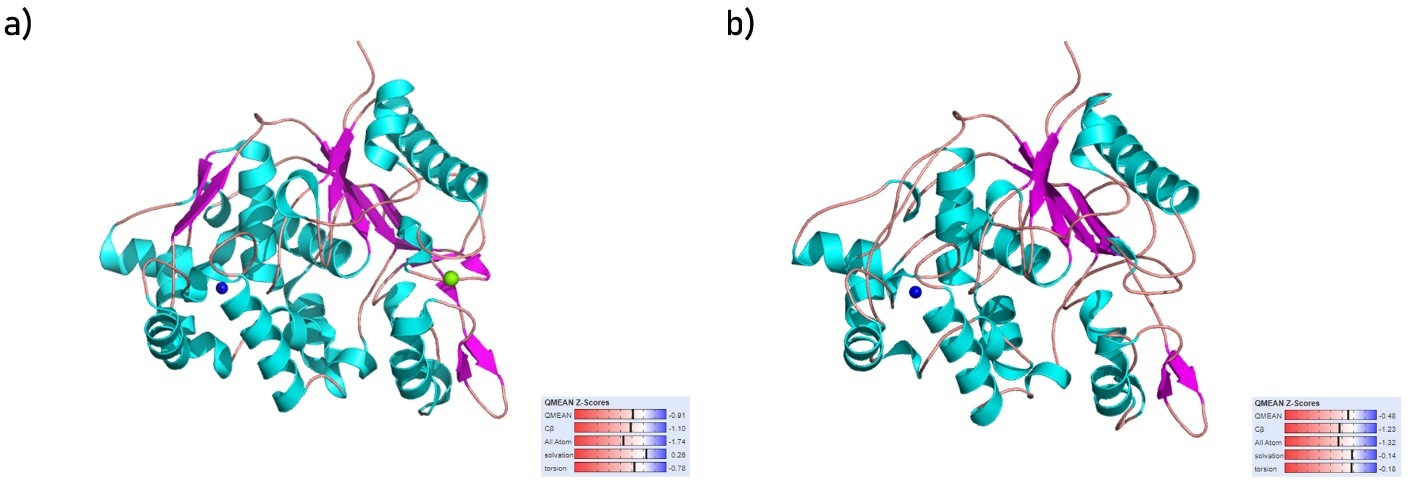
The structures of the Staphylococcus aureus lipase enzyme (Fig. 1a), and Staphylococcus warneri lipase enzyme (Fig. 1b) were successfully modeled by homology modeling. Fig. 1 displays the modeled structures along with their QMEAN Z-Scores, a scale used to evaluate the quality of a predicted structure (absolute zero indicates a perfect structure and beyond –4.0 indicates low-quality prediction).
|
Fig. 1. Structure of (a) Staphylococcus aureus lipase enzyme and (b) Staphylococcus warneri lipase enzyme along with their QMEAN Z-scores. Cartoon representation of protein structure colored based on their secondary structure (cyan for α‑helices, magenta for β-sheets, and salmon for loop regions), blue and green beads represent Zn and Ca cofactors respectively |
The top 3 ligands with the highest affinity for Staphylococcus aureus lipase enzyme were compounds a1970, a1883, and a1310 with binding affinities of –11.90, –11.90, and –11.80 kcal · mol–1, respectively. The top 3 ligands with the highest affinity for Staphylococcus warneri lipase enzyme were compounds w0855, w1914, and w1734 with binding affinities of –11.40, –11.10, and –10.80 kcal · mol–1, respectively.
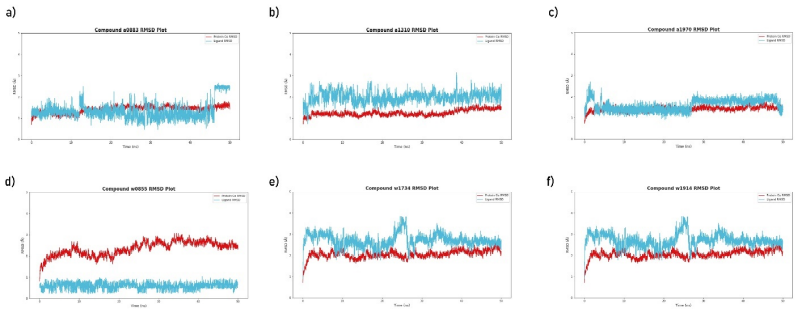
The RMSD plots for the top 3 ligands that were simulated with the Staphylococcus aureus lipase enzyme are shown in Fig. 2 (a-c) and the RMSD plots for the top 3 ligands that were simulated with the Staphylococcus warneri lipase enzyme is shown in Fig. 2 (d-f). The red lines represent the RMSD of the enzyme’s Cα (i.e., movement of the proteins backbone with respect to its initial position) and the cyan lines represent the RMSD of the respective ligands (i.e., movement of the ligands from their docked position).
|
Fig. 2. The RMSD plot for each of the top 3 ligands for Staphylococcus aureus lipase enzyme (a-c) and the RMSD plot for each of the top 3 ligands for Staphylococcus warneri lipase enzyme (d-f) throughout the production simulation of 50 ns. The RMSD was calculated using the initial position of the enzyme and their respective ligand as the reference points. The red line indicates the RMSD of the enzyme’s Cα and the cyan lines indicate the RMSD of the respective ligand in the simulation. The data was generated via VMD (v1.9.3) and plots were visualized via matplotlib and seaborn libraries in python [Humphrey et al. 1996Humphrey, W., Dalke, A., Schulten, K. (1996). VMD: visual molecular dynamics. J. Mol. Graph., 14(1), 27–28, 33–38. https://doi.org/10.1016/0263-7855(96)00018-5, Hunter 2007Hunter, J.D. (2007). Matplotlib: A 2D graphics environment. Comput. Sci. Eng., 9(3), 90–95. https://doi.org/10.1109/MCSE.2007.55, Waskom 2021Waskom, M.L. (2021). seaborn: statistical data visualization. J. Open Source Softw., 6(60), 3021. https://doi.org/10.21105/joss.03021] |
Compounds a0883 and a1970 simulated with the Staphylococcus aureus lipase enzyme generated stable RMSD plots throughout the production simulation (cyan lines in Fig. 2 (a and c) deviates < 2 Å) indicating their potential inhibitory activity, Fig. 3 (a) visualizes the biomolecular interactions compound a0883 forms the Staphylococcus aureus lipase enzyme and Fig. 3 (b) visualizes the biomolecular interactions compound a1970 forms the Staphylococcus aureus lipase enzyme. Compound w0855 simulated with the Staphylococcus warneri lipase enzyme generated stable RMSD plots throughout the production simulation (cyan lines in Fig. 2 (d) deviates < 2 Å) indicating their potential inhibitory activity, Fig. 3 (c) visualizes the biomolecular interactions compound w0855 forms the Staphylococcus warneri lipase enzyme. A brief summary of each of the lead compounds is given in Table 1.
|
Fig. 3. Biomolecular interactions between lead compounds and their respective lipase enzymes. (a) shows the interactions between compound a0883 with Staphylococcus aureus lipase enzyme, (b) shows the interactions between compound a1970 with Staphylococcus aureus lipase enzyme, and (c) shows the interactions between compound w0855 with Staphylococcus warneri lipase enzyme. The lead compounds are shown as green sticks, the interacting amino acid residues are shown as yellow sticks. Hydrogen bonds are shown as magenta lines and hydrophobic interactions are shown as orange dashes. All measurements are in Å unit. |
Table 1. A brief summary of the top 3 lead compounds as depicted in Fig. 3 |
|||
Serial code |
PubChem CID |
Common name |
2D chemical structure |
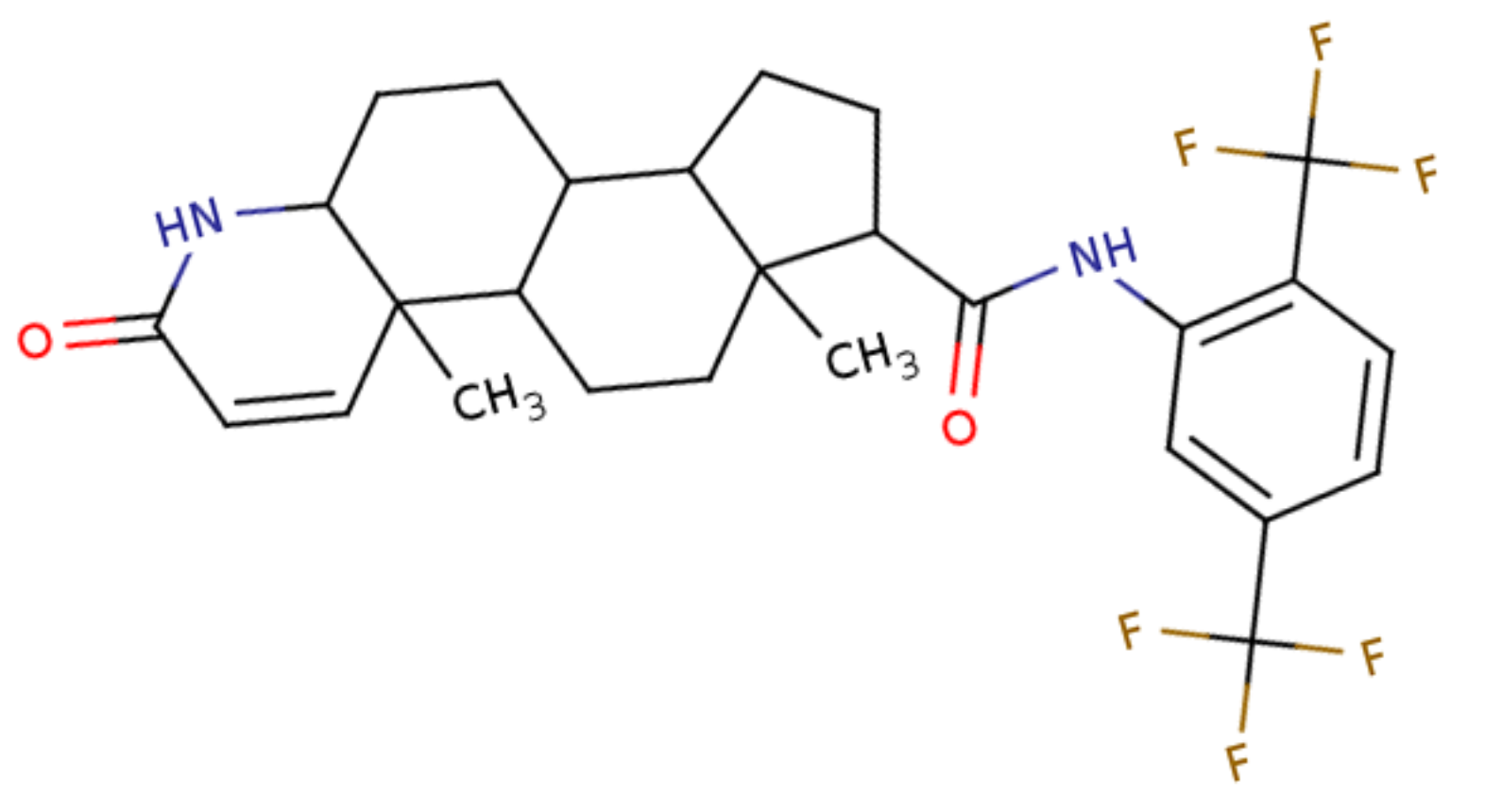
a0883 |
6918296 |
Dutasteride |
|
a1970 |
25176014 |
Entrectinib metabolite M5 |
|
w0855 |
68873 |
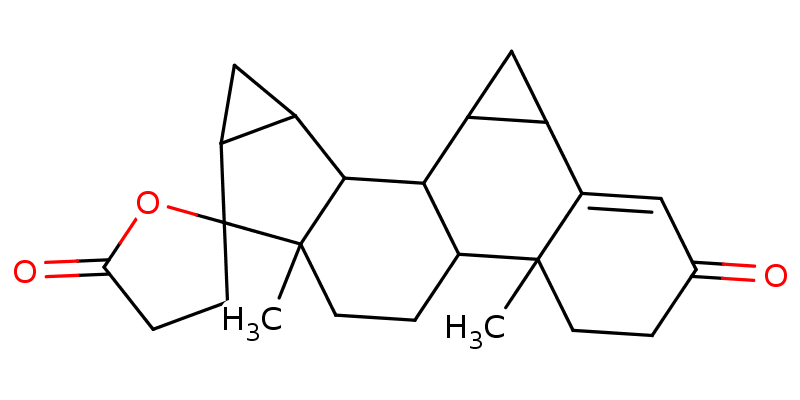
Drospirenone |
|
The interactions visualized in Fig. 3 are summarized in Table 2, Dutasteride forms 5 hydrophobic interactions and 2 hydrogen bonds with Staphylococcus aureus lipase, and Entrectinib metabolite M5 forms 4 hydrophobic interactions and 2 hydrogen bonds with the same enzyme. Drospirenone forms 10 hydrophobic interactions and 2 hydrogen bonds with Staphylococcus warneri lipase.
Table 2. Summary of the interactions between the top 3 lead compounds with their respective lipase enzyme |
|||
Interacting pair |
Interacting residue |
Distance (Å) |
Interaction type |
Dutasteride with Staphylococcus aureus lipase |
PHE17 |
3.67 |
Hydrophobic interaction |
PHE17 |
3.71 |
Hydrophobic interaction |
|
LEU18 |
3.22 |
Hydrophobic interaction |
|
PHE285 |
3.47 |
Hydrophobic interaction |
|
VAL350 |
3.44 |
Hydrophobic interaction |
|
PHE17 |
3.62 |
Hydrogen bond |
|
HIS349 |
2.94 |
Hydrogen bond |
|
Entrectinib metabolite M5 with Staphylococcus aureus lipase |
ALA174 |
3.87 |
Hydrophobic interaction |
PHE178 |
3.82 |
Hydrophobic interaction |
|
LEU242 |
3.95 |
Hydrophobic interaction |
|
VAL309 |
3.95 |
Hydrophobic interaction |
|
PHE17 |
3.48 |
Hydrogen bond |
|
SER116 |
4.03 |
Hydrogen bond |
|
Drospirenone with Staphylococcus warneri lipase |
ASN411 |
3.17 |
Hydrophobic interaction |
TYR412 |
3.72 |
Hydrophobic interaction |
|
TYR412 |
3.40 |
Hydrophobic interaction |
|
TYR437 |
3.33 |
Hydrophobic interaction |
|
LEU473 |
3.57 |
Hydrophobic interaction |
|
LEU473 |
3.74 |
Hydrophobic interaction |
|
LEU473 |
3.42 |
Hydrophobic interaction |
|
GLU484 |
3.75 |
Hydrophobic interaction |
|
LEU599 |
3.56 |
Hydrophobic interaction |
|
HIS602 |
3.60 |
Hydrophobic interaction |
|
GLU485 |
3.58 |
Hydrogen bond |
|
LEU599 |
3.99 |
Hydrogen bond |
|
Table 3 summarizes the averaged binding free energy for each lead compound against their respective lipase enzyme. The binding free energies calculated by the PRODIGY algorithm for Dutasteride, Entrectinib metabolite M5, and Drospirenone against their corresponding lipase enzymes were –12.84, –10.85, and –10.54 kcal · mol–1, respectively. These values were averaged with their corresponding binding affinities calculated by AutoDock Vina during the molecular docking step [Trott and Olson 2010Trott, O., Olson, A. J. (2010). AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem., 31(2), 455–461. https://doi.org/10.1002/jcc.21334].
Table 3. Average free binding energy calculation for each lead-lipase pair (all energy terms are in kcal Å · mol–1) |
|||
Interacting pair |
ΔGDocking |
ΔGPRODIGY |
ΔGAverage* |
Dutasteride with Staphylococcus aureus lipase |
–11.90 |
–12.84 |
–12.37 |
Entrectinib metabolite M5 with Staphylococcus aureus lipase |
–11.90 |
–10.85 |
–11.38 |
Drospirenone with Staphylococcus warneri lipase |
–11.40 |
–10.54 |
–10.97 |
*ΔGAverage was calculated via ΔGAverage = (ΔGDocking + ΔGPRODIGY)/2
Mastitis is known as the most expensive disease among cattle in the dairy industry costing farmers billions in lost revenues and the most common cause for its prevalence is known to be environmental pathogens such as Streptococcus spp., Enterobacteriaceae, or Staphylococcus spp. [Gill et al. 1990Gill, R., Howard, W.H., Leslie, K.E., Lissemore, K. (1990). Economics of Mastitis Control. J. Dairy Sci., 73(11), 3340–3348. https://doi.org/10.3168/jds.S0022-0302(90)79029-7, Chen and Alonzo 2019Chen, X., Alonzo, F. (2019). Bacterial lipolysis of immune-activating ligands promotes evasion of innate defenses. Proc. Natl. Acad. Sci., 116(9), 3764–3773. https://doi.org/10.1073/pnas.1817248116]. Elevation of SCC in milk is one of the most common indicators used for the assessment of mastitis risk in cattle and to no surprise, the infections caused by the former pathogens are known to elevate the SCC in cattle milk [Bianchi et al. 2019Bianchi, R.M., Schwertz, C.I., de Cecco, B.S., Panziera, W., De Lorenzo, C., Heck, L.C., Snel, G.G.M., Lopes, B.C., da Silva, F.S., Pavarini, S P., Driemeier, D. (2019). Pathological and microbiological characterization of mastitis in dairy cows. Trop. Anim. Health Prod., 51(7), 2057–2066. https://doi.org/10.1007/s11250-019-01907-0, Puppel et al. 2020Puppel, K., Kalińska, A., Kot, M., Slósarz, J., Kunowska-Slósarz, M., Grodkowski, G., Kuczyńska, B., Solarczyk, P., Przysucha, T., Gołębiewski, M. (2020). The Effect of Staphylococcus spp., Streptococcus spp. and Enterobacteriaceae on the Development of Whey Protein Levels and Oxidative Stress Markers in Cows with Diagnosed Mastitis. Anim. an Open Access J. from MDPI, 10(9), 1591. https://doi.org/10.3390/ani10091591]. By far the most common method used by farmers around the globe to combat mastitis is the use of antibiotics, which could be an efficient treatment to cure the infections by such pathogens, however, prolonged use of such antibiotics and their usage in prevention rather than treatment raise the growing issue of antibiotic-resistant bacteria and even the bioconcentration of such therapeutics in the cattle’s milk and meat [Pyörälä 2002Pyörälä, S. (2002). New Strategies to Prevent Mastitis. Reprod. Domest. Anim., 37(4), 211–216. https://doi.org/10.1046/j.1439-0531.2002.00378.x, Shim et al. 2004Shim, E.H., Shanks, R.D., Morin, D.E. (2004). Milk Loss and Treatment Costs Associated with Two Treatment Protocols for Clinical Mastitis in Dairy Cows. J. Dairy Sci., 87(8), 2702–2708. https://doi.org/10.3168/jds.S0022-0302(04)73397-4, Ebrahimi et al. 2019Ebrahimi, M., Mohammadi-Dehcheshmeh, M., Ebrahimie, E., Petrovski, K.R. (2019). Comprehensive analysis of machine learning models for prediction of sub-clinical mastitis: Deep Learning and Gradient-Boosted Trees outperform other models. Comput. Biol. Med., 114, 103456. https://doi.org/10.1016/j.compbiomed.2019.103456]. Among the recent studies that explored the mechanism in which several pathogens successfully evaded the cattle’s immune system, it was observed that certain pathogens utilized their lipase enzymes to hydrolyze the bacterial-derived lipoproteins (lipolysis), foreign lipids (and/or lipoproteins) being an inducer for immune response, their hydrolyzation offers the pathogen’s the ability to evade the host's immune response and survive within the cattle’s milk [Chen and Alonzo 2019Chen, X., Alonzo, F. (2019). Bacterial lipolysis of immune-activating ligands promotes evasion of innate defenses. Proc. Natl. Acad. Sci., 116(9), 3764–3773. https://doi.org/10.1073/pnas.1817248116]. Therefore, the inhibition or disruption of these lipase enzymes could potentially counteract the evasion mechanism of such pathogens and provide the upper hand for the cattle’s immune response to detect and eradicate such pathogens more efficiently without the need to administer antibiotics.
In this study, computational drug discovery (also known as computer-aided drug discovery, CADD) was exploited to repurpose already FDA-approved drugs (for humans) for use in cattle, while the physiology of animals may differ to some extent to those of animals, drugs deemed safe for human consumption are likely to perform more efficiently in in vitro and in vivo that their novel counterparts as their toxicological side effects are already extensively researched. A common example for such drugs is NSAIDs (Non-steroidal anti-inflammatory drugs) which are FDA-approved for human use but are also administered in veterinary medicine for the same purpose [Modi et al. 2012Modi, C., Kumar, A., Avale, M. (2012). Toxicopathological overview of analgesic and anti-inflammatory drugs in animals. J. Appl. Pharm. Sci., 02(01), 149–157. Google Scholar, Park 2019Park, K. (2019). A review of computational drug repurposing. Tcp, 27(2), 59–63. https://doi.org/10.12793/tcp.2019.27.2.59].
Staphylococcus aureus and Staphylococcus warneri being among the most common cattle pathogens, their respective enzymes were selected as a target for repurposing of FDA-approved drugs as novel inhibitors. The sequence for the lip1 and lipWY genes of Staphylococcus aureus and Staphylococcus warneri respectively was retrieved from the Uniprot database and their corresponding 3D structure was modeled via homology modeling. As shown in Fig. 1, both the enzymes were modeled with above-average QMEAN Z-scores (dashes closer to the blue regions indicate more accurate structures). The virtual screening (molecular docking) experiment with each of the 2 lipases against the library of 1993 FDA-approved drugs yielded several promising hits, with the top 3 hits for each lipase scoring beyond –11.00 kcal · mol–1, however, molecular docking alone has its limitation as it neglects the physiological cellular environment and considers mainly the geometrical optimization of the protein and ligand in its scoring, hence, cross-validation of such experiments are critical [Huang and Zou 2010Huang, S.-Y., Zou, X. (2010). Advances and challenges in protein-ligand docking. Int. J. Mol. Sci., 11(8), 3016–3034. https://doi.org/10.3390/ijms11083016].
Molecular dynamics on the other hand is among the most sophisticated methods used in computational drug discovery which provides nanoscale insights into the atom-level interactions between a protein and its ligand [Borhani and Shaw 2012Borhani, D.W., Shaw, D.E. (2012). The future of molecular dynamics simulations in drug discovery. J. Comput. Aided. Mol. Des., 26(1), 15–26. https://doi.org/10.1007/s10822-011-9517-y, De Vivo et al. 2016De Vivo, M., Masetti, M., Bottegoni, G., Cavalli, A. (2016). Role of Molecular Dynamics and Related Methods in Drug Discovery. J. Med. Chem., 59(9), 4035–4061. https://doi.org/10.1021/acs.jmedchem.5b01684]. The 6 pairs of lipases with their corresponding hit compounds simulated under physiological conditions (0.15 mol · L–1 salt and 310.15 K) yielded promising results only for 3 of the 6 hit compounds. The plots in Fig. 2 summarized the motion of the lipase enzyme and corresponding hit compound, among the hit compounds simulated with the Staphylococcus aureus lipase enzyme, only compound a0883 (Fig. 2a) and a1970 (Fig. 2b) maintained close proximity to the lipase enzyme throughout the simulation period (the cyan line which represents the motion of the ligand, deviates less than 2 Å throughout the simulation, this indicates its preference and stability of its initial docked pose). However, the spike of compound a0883 (Fig. 2a) beyond the 2 Å threshold at the ending of the simulation indicates potential inconclusiveness as such deviation could indicate drifting from the docked pose, likewise, compound a1970 made an initial spike above the 2 Å threshold at the beginning of the simulation by remained stable after that. Compound w0855 that was simulated with Staphylococcus warneri lipase enzyme yielded the most promising results by deviating less than 1 Å throughout the simulation indicating its potential strong affinity (inhibitory potential) to the Staphylococcus warneri lipase enzyme.
The promising results of compound w0855 can further be confirmed by the interaction profiles it makes with the Staphylococcus warneri lipase enzyme, as shown in Fig. 3 (c) and Table 2, it forms more hydrophobic interactions (10 in total) with the former enzyme along with 2 hydrogen bonds that further stabilize its position. Compounds a0883 and a1970 on the other hand make half of the hydrophobic interactions that compound w0855 made. Considering the binding free energies that were estimated from the PRODIGY sever and AutoDock Vina, all of the 3 lead compounds tend to be exothermic in their association with their respective enzymes, moreover, the ΔG calculated by both of AutoDock Vina and PRODIGY sever are close to each other, reducing the overall estimation bias.
Considering the computational analysis performed in this study and the criteria applied for lead selection, compound w0855, commonly known as Drospirenone, exhibits potential lipase inhibitory activity against the lipWY lipase of Staphylococcus warneri. Drospirenone is a unique progestogen with anti-mineralocorticoid properties, it is one of several different progestins that are commonly used in birth control pills and in menopausal hormone therapy [Oelkers 2004Oelkers, W. (2004). Drospirenone, a progestogen with antimineralocorticoid properties: a short review. Mol. Cell. Endocrinol., 217(1), 255–261. https://doi.org/10.1016/j.mce.2003.10.030]. Given its common use worldwide, the risk of it possessing a toxic effect on cattle physiology can be quite low, however, its efficiency and efficacy as an anti-microbial agent and its effect on the cattle’s reproduction cycle (and/or offspring health) can only be explained by further in vitro and in vivo assays. Moreover, this research can also serve as a precursor for the use of computational drug discovery approaches to repurpose drugs that already have sufficient data regarding their toxicity, efficacy, and efficiency to combat pathogens affecting the dairy industry.
Mastitis is the single most expensive disease in the dairy industry and the most common combat mechanism used by farmers around the globe is the administration of antibiotics (and probiotics). Antibiotics can be the most accessible and safest solution to combat mastitis, however, it’s long-term use comes with many disadvantages. In this paper, computational drug repurposing study was performed to find hit compounds that could inhibit the lipase activity of specific pathogens known to cause mastitis in cattle among a dataset of already FDA-approved drugs. Based on the computational analyses performed in this paper and criteria applied for lead selection, Drospirenone, a unique progestogen with anti-mineralocorticoid properties, commonly used in birth control pills was concluded to exhibit a potential lipase activity inhibitor against the lipWY lipase of Staphylococcus warneri.
The computational calculations reported in this paper were performed on TUBITAK ULAKBIM, High Performance and Grid Computing Center (TRUBA resources). This research did not receive any monetary funding.
Received: 30 Oct 2021
Accepted: 8 Jan 2022
Published online: 7 Apr 2022
Accesses: 929
Ghafoor, N.A., Sitkowska, B., (2021). Computational repurposing of FDA-approved drugs against specific mastitis-causing pathogens. Acta Sci. Pol. Zootechnica, 20(4), 5–14. DOI: 10.21005/asp.2021.20.4.02.