Review Article
Piotr Guliński ![]()
Institute of Animal Production and Fisheries, Siedlce University of Natural Sciences and Humanities, 08-110 Siedlce, ul. Prusa 14, Poland
Abstract. Domestic cattle descend from the aurochs, a species of wild animals, inhabiting forests of Europe and Asia in the past. The results of genetic studies allow concluding that the world cattle population originate from 80 female aurochs domesticated in today's southeastern Turkey. After 10,500 years the worldwide bovine population has exceeded 1.4 billion, with two subspecies: taurus cattle (Bos taurus) and zebu cattle (Bos indicus). The former was first scientifically described by Linnaeus, who in 1758 proposed its name, used until now. The processes of cattle domestication and improvement have led to more than 1000 distinct breeds. During the last two centuries several kinds of classification have been developed in order to identify cattle breeds. Several criteria have been used, such as basic cranial type, length of the horns, skull shape, degree of domestication, production purpose, type of ancestral aurochs, geographic distribution, place of breed origin, population size, growth performance, biochemical polymorphisms and molecular-genetic methods. At present the attention of cattle farmers around the world has focused on the most productive improved breeds at the expense of less productive local ones. Modern methods, such as artificial insemination and embryo transfer, have allowed unlimited spread of breeding material around the world. This way, it has become possible to use reproductive material of different cattle breeds on an unprecedented scale in many countries. At present, many of the latter have disappeared, or their population is on the verge of extinction.
Keywords: cattle, breeds, origin, criteria for classification
The wild ancestor of modern cows was the aurochs (Bos primigenius, Bojanus 1827). Aurochs lived in Asia, Europe, and North Africa, but they were domesticated in the Fertile Crescent region of the Middle East between 10,300 and 10,800 years ago. This took place in what is now south-western Turkey, near its border with Syria [Felius 1995Felius, M. (1995). Cattle Breeds: An Encyclopedia. Trafalgar Square Publishing. Google Scholar]. According to Zeder [2012]Zeder, M.A. (2012). Pathways to animal domestication in biodiversity in agriculture: Domestication, Evolution, and Sustainability. Eds. P. Gepts, T.R. Famula, R.L. Bettingeret, et al., Cambridge University Press. Google Scholar, the so-called prey pathway was used in the domestication of cattle and other large and medium-sized herbivores. Archaeological evidence from the Middle East suggests that Bos taurus followed this pathway to domestication, with change from generalized to specialized hunting and subsequent herd management in the natural habitats of its wild ancestor [Zeder 2009Zeder, M.A. (2009). The Neolithic macro-(r)evolution: Macro evolutionary theory and the study of culture change. J. Archeol. Res., 171, 1–63. https://doi.org/10.1007/s10814-008-9025-3, Zeder 2011Zeder, M.A. (2011). The origins of agriculture in the Near East. Curr. Anthropol., 52(S4), 221–225. https://doi.org/10.1086/659307]. Domesticated aurochs then developed into humpless domestic cattle (Bos taurus). Later, from the Fertile Crescent they spread to Eurasia and Africa. The second place of aurochs domestication was the Indian subcontinent, i.e. modern India and Pakistan. About 2,000 years after the first domestication, the next took place in the Indus Valley, with an aurochs subspecies later evolving into Bos indicus [Patel 2009Patel, A. (2009). Occupational histories, settlements, and subsistence in Western India: what bones and genes can tell us about the origins and spread of pastoralism. Anthropozool., 44, 173–188. https://doi.org/10.5252/az2009n1a8]. The theory is that western Egypt was the third place where aurochs were domesticated supposedly about 9,000 years ago, which is highly questioned in the light of recent research results [Pitt et al. 2018Pitt, D., Sevane, N., Nicolazzi, E.L., MacHugh, D.E., Park, S., Colli, L., Martinez, R., Bruford, M.W., Orozco-terWengel, P. (2018). Domestication of cattle: Two or three events? Evol. Appl., 12(1), 123–136. https://doi.org/10.1111/eva.12674].
The original scientific name of this animal – Bos primigenius – was translated into German as Auerochse or Urochs, literally meaning “primary ox” or “proto ox”. Aurochs appeared on Earth about two million years ago in the area where India is now [Scheu et al. 2015Scheu, A., Powell, A., Bollongino, R., Vigne, J.D., Tresset, A., Çakırlar, C., Benecke, N., Burger J. (2015). The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet., 16–54. https://doi.org/10.1186/s12863-015-0203-2]. Later, about 250,000 years ago, it spread to present-day Europe. It is believed that there were three subspecies – Bos primigenius primigenius in Europe and the Middle East, Bos primigenius nomadicus in India and Bos primigenius mauretanicus in North Africa. Aurochs are among the famous animal images in Paleolithic cave paintings, such as those found in 1940 in Lascaux [Ajmone-Marsha et al. 2010Ajmone-Marsha, P., Garcia, F.J., Lenstra, J.A. (2010). On the origin of cattle: how aurochs became cattle and colonized the world. Evol. Anthropol., 19, 148–157. https://doi.org/10.1002/evan.20267], France or those found in 1868 in Altamira, Spain [Cuenca-Solana et al. 2016Cuenca-Solana, D., Gutierrez-Zugasti, I., Ruiz-Redondo, A., Gonzalez-Morales, M.R., Setien, J., Ruiz-Martínez, E., Palacio-Perez, E., de las Heras-Martín, C., Prada-Freixedo, A., Lasheras-Corruchaga, J.A. (2016). Painting Altamira Cave? Shell tools for ochre-processing in the Upper Palaeolithic in northern Iberia. J. Archaeol. Sci., 74, 135–151. https://doi.org/10.1016/j.jas.2016.07.018]. Radiocarbon dating has shown that these paintings are 17–19 thousand years old [Leroi-Gourhan 1982Leroi-Gourhan, A. (1982). The Archaeology of Lascaux Cave. Sci. Am., 246(6), 104–113 https://doi.org/10.1038/scientificamerican0682-104, Laval 2007Laval, F. (2007). Mon père, l'Homme de Lascaux. Pilot 24, France. Google Scholar]. In Lascaux, the Hall of the Bulls is particularly interesting, with the walls covered with large-sized (4–5-meter-long) silhouettes of bulls with black outlines and filled with spots of red and black. Additionally, the majority of 179 petroglyphs (76%) found in 2007 in Qurta, Egypt, on the eastern bank of the Nile, depict wild aurochs. The largest figure found there is about 1.8 meters long. Drawings were created 15,000 years ago and are assessed by archaeologists as the oldest evidence of human material activity in Egypt. This place is also called Lascaux on the Nile [Huyge and Claes 2008Huyge, D., Claes, W. (2008). `Ice Age' art along the Nil. Egyptian Archaeology. Bull. Egypt Expl. Soc., 33, 25–28. Google Scholar].
In the 1950s, a Neolithic human settlement from 7400 to 7300 BC was discovered in Çatal Hüyük, Turkey. It has been proven that those people were engaged in cattle and sheep farming and plant cultivation. The murals discovered in this settlement, with figurines and art installations made, among others, from the heads of bulls demonstrate that those animals played a key role in the lives of those people [Twiss and Russel 2009Twiss, K., Russel, N. (2009). Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük (Turkey). Paléorient 35(2), 19–32. https://doi.org/10.3406/paleo.2009.5296].
Cave and other paintings, literary descriptions [Gajusz Pliniusz Sekundus 2019Gajusz Pliniusz Sekundus (2019). Historia Naturalna. Tom II. Antropologia i zoologia. Księgi VII–XI [Natural History. Volume II. Anthropology and Zoology. Books VII–XI]. Wydaw. Nauk. UMK, Toruń [in Polish]. Google Scholar] and material evidence, like skeletons, indicate that aurochs were giant animals. The males were almost 2 m high at the withers and up to 2.5 m together with the horns. In aurochs, as in other primitive animals, sexual dimorphism was very sharply outlined. Females reached up to 1.4–1.5 m in height at the withers. Brown-black males had a bright stripe along their back. Brown cows were smaller and, together with red calves, lived in large herds. Aurochs lived up to 25 years, forming loose groups consisting of adult females, calves, and young males and females. Adult males usually lived alone. Aurochs adapted perfectly to different environmental conditions. Their habitat was forests and woods from the Middle East through continental Europe and even to the cold climate of the British Isles. Aurochs made great use of lush vegetation, but, unfortunately, they were also an ideal target for people who hunted them for meat as well as for recreation. Since ancient times, aurochs had impressed people with their greatness, courage, and strength. The life force and vitality of these animals had been admired by many generations.
In Anatolia and the Middle East, aurochs were worshipped and considered sacred animals [Twiss and Russel 2009Twiss, K., Russel, N. (2009). Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük (Turkey). Paléorient 35(2), 19–32. https://doi.org/10.3406/paleo.2009.5296]. Despite the early extinction of their contemporaries, i.e. the mammoth and saber-toothed tiger, they managed to survive an astonishingly long time. In Europe, west of the Oder, the aurochs was wiped out at the turn of the 12th century. It survived only in central Europe: Poland, Lithuania and East Prussia.
From the extant records of diplomats residing in Poland in the 14th century, it is known that Polish princes and kings had been the owners of all aurochs at least since 1288. Polish rulers protected their last herd living in the forests near Sochaczew, which effectively extended the existence of the species by several centuries. The need for such protection was understood by King Jagiełło, and it was continued by all the successors of the Jagiellonian dynasty, and later also by the House of Vasa. The last collective hunt for these animals was carried out in 1410 [Długosz 2020Długosz, J. (2020). Roczniki, czyli kroniki sławnego Królestwa Polskiego [Annals, or chronicles of the famous Kingdom of Poland]. Księga 10 i 11. Wyd. Nauk. PWN Warszawa. Wydanie II [in Polish]. Google Scholar]. As aurochs population had steadily declined, hunting for these animals was banned and punished by death. In 1564, only 38 Polish animals of this species survived. The last aurochs, an adult female [Daszkiewicz and Edel 2014Daszkiewicz, P., Edel, P. (2014). The will of Ludwig Heinrich Bojanus (1776–1827), an interesting nineteenth-century natural history document. Arch. Nat. Hist., 41, 164–167. https://doi.org/10.3366/anh.2014.0221], died in Poland in the Kampinos Forest in 1627 [Frisch 2010Frisch, W. (2010). Der Auerochse – das europäische Rind. Starnberg, Watzmannstr. 4 I. Google Scholar]. The causes of aurochs extinction were most likely excessive hunting, a reduction in the pasture area grazed by domesticated cattle, and a growing lack of genetic diversity leading to their overall weakening and reducing their resistance to disease.
The first scientific description of aurochs based on its skeleton was made in 1827 by Ludwig Henryk Bojanus, a Polish professor at Stefan Batory University in Vilnius. He used the name Bos primigenius referring to this species, to which `Bojanus, 1827' was added later [Bojanus 1965Bojanus, L.H. (1965). De uro nostrate eiusque sceleto commentatio = Rozważania nad żubrem i jego kośćcem, 1825. W 140-lecie krytycznie opracowali i komentarzem opatrzyli Tadeusz Roskosz i Wojciech Empel [\emph{De uro nostrate eiusque sceleto commentatio} = Discourse on the bison and its skeleton, 1825. On the 140th anniversary, Tadeusz Roskosz and Wojciech Empel provided a critical review and commentary]. Memorabilia Zoologica, 14 [in Polish]. Google Scholar]. He is the author of a 1825 work De uro nostrate eiusque sceleto commentatio (Reflections on the bison and its skeleton), in which, on the basis of a meticulous analysis of the skeletons of both the aurochs and the European bison, he proved that they were two distinct species.
Bones found in Neolithic ceramics in Argissa-Magula, Thesis, Greece, constitute the earliest evidence of domesticated cattle in Europe [Conolly et al. 2012Conolly, J., Manning, K., Colledge, S., Dobney, K., Shennan, S. (2012). Species distribution modelling of ancient cattle from early Neolithic sites in SW Asia and Europe. Holocene, 24, 3–14. https://doi.org/10.1177/0959683612437871]. Their age is estimated as 8,500 years. Remains of the first European farms suggest two routes for the migration of domesticated cattle: along the Mediterranean coast and along the Danube. Following the first route, about 7,900–7,700 years ago the cattle reached southwestern France, Corsica, Languedoc and the east coast of Spain [Gkiasta et al. 2003Gkiasta, M., Russell, T., Shennan, S., Steele, J. (2003). Neolithic transition in Europe: The radiocarbon record revisited. Antiquity, 77, 45–62. https://doi.org/10.1017/S0003598X00061330, Cymbron et al. 2005Cymbron, T., Freeman, A., Malheiro, M.I., Vigne, J.D., Bradley, D. (2005). Microsatellite diversity suggests different histories for Mediterranean and Northern European cattle populations. Proc. R. Soc. Lond. B Biol. Sci., 272, 1837–1843. https://doi.org/10.1098/rspb.2005.3138, Tresset and Vigne 2007Tresset, A., Vigne, J.D. (2007). Substitution of species, techniques and symbols at amine transporter, causes complex vertebral malformation. Proc. Br. Acad., 144, 189–210. Google Scholar], and about 1,000 years later along the Danube they moved to Central and Northern Europe.
By analyzing degraded fat residue on archaeological sites, the researchers found out that Neolithic farmers living in the UK and Northern Europe might have been among the first to start producing milk for human consumption. Recently, isotopic analysis of pottery residues has allowed the differentiation between meat and dairy fats [Copley et al. 2003Copley, M.S., Berstan, R., Dudd, S.N., Docherty, G., Mukherjee, A.J., Straker, V., Payne, S., Evershed, R.P. (2003). Direct chemical evidence for widespread dairying in prehistoric Britain. Proc. Natl. Acad. Sci. USA, 100, 1524–1529. https://doi.org/10.1073/pnas.0335955100, Evershed et al. 2008Evershed, R.P., Payne, S., Sherratt, A.G., Copley, M.S., Coolidge, J., Urem-Kotsu, D., Kotsakis, K., Özdoğan, M., Özdoğan, A.E., Nieuwenhuyse, O., Akkermans, P., Bailey, D., Radian-Romus Andeescu, R-R., Campbell, S., Farid, S., Hodder, I., Yalman, N., Özbaşaran, M., Bıçakcı, E., Garfinkel, Y., Thomas Levy, T., Burton M.M. (2008). Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding. Nature, 455, 528–531. https://doi.org/10.1038/nature07180] and has indicated that milk was being utilised since the early Neolithic. Additionally, the variation in the nitrogen isotope ratios in Neolithic cattle teeth suggests that calves were weaned early [Balasse and Tresset 2002Balasse, M., Tresset, A. (2002). Early weaning of Neolithic domestic cattle (Bercy, France) revealed by intra-tooth variation in nitrogen isotope ratios. J. Archaeol. Sci., 29, 853–859. https://doi.org/10.1006/jasc.2001.0725] making milk available for human consumption.
According to the researchers, the full capacity to digest milk (lactose) developed in this population between 5,000 and 4,000 BC, owing to a genetic mutation that made it possible to produce the lactase enzyme. If the above dates were to be accurate, the emergence of other great milk-drinking civilizations in the Middle East, India, and North Africa must have taken place at the same time. Recent studies indicate that the ability of European population to digest milk sugar had developed first in farming communities in the central part of the continent and not, as previously thought, in human groups living in northern regions. According to those studies, genetic changes that allowed Europeans to drink milk without health consequences began 7,500 years ago in the Balkans [Leonardi et al. 2012Leonardi, M., Gerbault, P., Thomas, M.G., Burger, J. (2012). The evolution of lactase persistence in Europe. A synthesis of archeological and genetic evidence. Int. Dairy J., 22, 88–97. https://doi.org/10.1016/j.idairyj.2011.10.010].
In the Babylonian civilization (2000 years BC), cattle were kept primarily as a source of draft power, not for milk. According to old Babylonian texts record, monetary value for draft ox was 12 shekels, compared with 7 ⅔ shekels for milking cow [Stol 1995Stol, M. (1995). Old Babylonian cattle. Bull. Sumer. Agric., 8, 173–213. https://ccp.yale.edu/old-babylonian-cattle. Google Scholar].
During the Hellenistic period (around 330–63 BC) in Greece, cattle were used for transport, ritual slaughter, and beef and milk production. According to Aristotle, the rich pastures of Epirus were famous for their large cattle production, with up to 30 l of milk per cow per day obtained [Aristotle 350 BCAristotle (350 BC). In Historia Animalium. Google ScholarAristotle (350 BC). In Historia Animalium. Google Scholar]. The remains found in Kassope in Epirus made it possible to determine the height of cattle in the 7th to 8th centuries BC at 115–135 cm at the withers [Kron 2002Kron, G. (2002). Archaeozoological evidence for the productivity of Roman livestock farming. Münstersche Beiträge zur Antiken Handelsgeschichte, 21(2), 53–73. Google Scholar]. It is also known that Epirus beef was exported to many regions of Italy and southern France. It is believed that the cattle were probably the ancestors of large Roman breeds.
Cattle were also raised during the Roman Empire [Fussel 1972Fussel, G.E. (1972). The Classical Tradition in West European Farming. Newton Abbot, Devon, UK. Google Scholar]. They were described in detail in the works of Cato, Varro, Columella, and Pliny [MacKinnon 2010MacKinnon, M. (2010). Cattle ``breed'' variation and improvement in Roman Italy: Connecting the zooarcheological and ancient textual evidence. World Archeol., 42, 55–73. https://doi.org/10.1080/00438240903429730]. In the Roman Empire, cattle were not milked, but they were used for transport and field work. Roman writers described the diversity of cattle in different regions of the Empire. Paleontological evidence indicates that in its different parts cows were very diverse in size. In southern Germany, for example, Germanic cattle reached a height of 95–125 cm at the withers, while in Roman provinces they were 100–150 cm tall. Pax Romana and the infrastructure of the Roman Empire facilitated the export of large Italian cattle breeds to remote provinces, where small indigenous cattle breeds were reared [Kron 2002Kron, G. (2002). Archaeozoological evidence for the productivity of Roman livestock farming. Münstersche Beiträge zur Antiken Handelsgeschichte, 21(2), 53–73. Google Scholar]. Interestingly, large cattle disappeared shortly after the fall of the Roman Empire, suggesting that smaller animals were better suited to breeding regression and to primitive technologies of their maintenance during the Middle Ages.
No definitive evidence for the domestication of indigenous cattle in China currently exists. The introduction of Bos taurus into that country occurred just prior to the development of the first Chinese States [Liu and Chen 2003Liu, L., Chen, X. (2003). State Formation in Early China. Duckworth London. Google Scholar]. Genetic and archaeological data both suggest that domesticated Bos taurus was introduced to China from West Asia between 3600 and 2000 BC [Barton and Cheng-Bang 2014Barton, L., Cheng-Bang, A. (2014). An evaluation of competing hypotheses for the early adoption o.f wheat in East Asia. World Archaeol., 46, 775–798. https://doi.org/10.1080/00438243.2014.953703, Liu et al. 2014Liu, X., Lightfoot, E., O'Connell, T. C., Wang, H., Li, S., Zhou, L., Hu, Y., Motuzaite-Matuzeviciute, G., Jones M. K. (2014). From necessity to choice: Dietary revolutions in West China in the second millennium BC. World Archeol., 46(5), 661–680. https://doi.org/10.1080/00438243.2014.953706, Jaang 2015Jaang, L. (2015). The landscape of China's participation in the Bronze Age Eurasian network. J. World Prehist., 28(3), 179–213. https://doi.org/10.1007/s10963-015-9088-2]. It is likely that Bos taurus entered China through the Gansu Qinghai region, but the exact timing and routes of introduction are still unknown, and it is unclear whether Bos taurus reached China as a single event or in multiple waves. Bos indicus probably entered southern China after 2000 BC, but the timing and routes of its introduction remain a mystery [Liu et al. 2014Liu, X., Lightfoot, E., O'Connell, T. C., Wang, H., Li, S., Zhou, L., Hu, Y., Motuzaite-Matuzeviciute, G., Jones M. K. (2014). From necessity to choice: Dietary revolutions in West China in the second millennium BC. World Archeol., 46(5), 661–680. https://doi.org/10.1080/00438243.2014.953706, Jaang 2015Jaang, L. (2015). The landscape of China's participation in the Bronze Age Eurasian network. J. World Prehist., 28(3), 179–213. https://doi.org/10.1007/s10963-015-9088-2].
The first cows were brought to America by Christopher Columbus. This happened during the second voyage in 1493 [Rouse 1977Rouse, J.E. (1977). The Criollo: Spanish Cattle in the Americas. University of Oklahoma Press, Norman, USA. Google Scholar, McTavish et al. 2013McTavish, E.J., Decker, J.E., Schnabel, R.D., Taylor, J.F., Hillis, D.M. (2013). New World cattle show ancestry from multiple independent domestication events. Proc. Natl. Acad. Sci. USA, 110(15), E1398-E1406. https://doi.org/10.1073/pnas.1303367110]. Over the next fifty years, every Spanish ship bound for America imported five or six young cattle, but it is assumed that only two or three of them survived. It is estimated that at most 300 Spanish cattle had reached America by this route. Cattle in the New World have evolved from both subspecies, i.e. both Zebu and European cattle [Felius et al. 2014Felius, M., Beerling, M.L., Buchanan, D.S., Theunissen, B., Koolmees, P.A., Lenstra, J.A. (2014). On the History of Cattle Genetic Resources. Diversity, 6(4), 705–750. https://doi.org/10.3390/d6040705]. In addition, historical records indicate that the natural selection of the oldest cattle breed in the U.S., i.e. Longhorns, is 450 years old, with about 80 to 200 generations [McTavish et al. 2013McTavish, E.J., Decker, J.E., Schnabel, R.D., Taylor, J.F., Hillis, D.M. (2013). New World cattle show ancestry from multiple independent domestication events. Proc. Natl. Acad. Sci. USA, 110(15), E1398-E1406. https://doi.org/10.1073/pnas.1303367110].
Commonly called `Hollander' cows, the Dutch Black-and-White cattle have been imported to Poland since the mid–16th century. The first known import of dairy cows from the Netherlands to Poland, by Mikołaj Firlej, the Castellan of Wiślice to his estate near Lubartów, took place as early as 1560. As a Calvinist, he provided shelter and protection to people persecuted in western Europe, especially those from the Netherlands and Flanders. They brought Frisian cows to their new place of residence and contributed as cattle breeders to the development of the material culture there [Kościszewska 1932Kościszewska, Z. (1932). Lewartów (Lubartów) w latach 1543– –1643 [Lewartów (Lubartów) in the years 1543–1643]. Pr. Sem. Hist. KUL, 1 [in Polish]. Google Scholar]. In the initial period, efforts to establish and run Hollander cattle breeding were undertaken by a few wealthy aristocrats, which meant the emergence of Polish Black-and-White cows. At the beginning of the 17th century, Black-and-White cows were very valuable; it is known that the price of such a cow at the time was 100 Polish zlotys and was more than six times higher than the price of indigenous grey cows [Wolski and Wisner 1969Wolski, K., Wisner, H. (1969). O krowy holenderskie pilno się staram [I am trying hard to get Dutch cows]. Kwart. Hist. Kult. Mat., 1 [in Polish]. Google Scholar]. There were other Dutch cattle breeders at that time in Poland. Among many others, Jan Zamoyski already reared Dutch cattle on his farm in Horyszów in 1597, in 1631 Lew Sapieha brought 26 such cows to his farm in Różana, and Krzysztof II Radziwiłł raised them in his estate in Birże [Wolski and Wisner 1969Wolski, K., Wisner, H. (1969). O krowy holenderskie pilno się staram [I am trying hard to get Dutch cows]. Kwart. Hist. Kult. Mat., 1 [in Polish]. Google Scholar]. Kluk [1779]Kluk, K. (1779). Zwierząt domowych i dzikich, osobliwie kraiowych, historyi naturalney początki i gospodarstwo. Tom 1. O zwierzętach ssących [Domestic and wild, particularly native animals: introduction to natural history and farming. Volume 1. On the suckling animals]. Drukarnia J. K. Mości i Rzeczypospolitej u Księży Pijarów, Warszawa [in Polish]. Google Scholar reports that in the second half of the 18th century, there were three main types of cattle in Poland: black and white (Dutch), red (so called `common'), and gray (Podolian).
Since Linnaeus, a species has been considered to be the primary systematic unit in the animal kingdom. It consists of a group of similar organisms with a common gene pool, being able to breed with one another. Organisms of the same species are of common origin, have a specific spatial distribution and cannot breed with other species. Thus, a species does not exchange its genes with another species due to its sexual isolation. Male interspecies hybrids are usually infertile, e.g. mules or the hybrid of the wisent and domestic cattle. Linnaeus defined species as a collection of similar individuals in the same way as the offspring are similar to their parents. Apart from introducing the concept of species, he formulated principles of naming them [Linne 1760Linne, C. (1760). Systema naturae per regna tria naturae secundum classes, ordines, genera, species cum characteribus, differentis, synonymis, locis, Tomus I. Praefatus est Joannes Ioachimus Langius, Halae Magderburgicae MDCCLX. https://doi.org/10.5962/bhl.title.7377]. Since then, the naming of species has been subject to binominal rules, i.e. using two words, in which the first part in the form of a noun is the generic name, identifying the genus, and the second is the so-called species epithet, that is to say, a species name, usually in the adjectival form. It should be noted that the species epithet for cattle (taurus) provided by Linnaeus refers to the Taurus Mountain range in southern Turkey, separating the Mediterranean coastal region from the central Anatolian Plateau. Binominal naming was first used in 1623 by Caspar Bauhin in Pinax Theatri Botanici [Bauhin 1623Bauhin, C. (1623). Pinax theatri botanici. Basileae Helvet. Sumtib us typis Ludovici Regis, MDCLXIII [in Latin]. Google Scholar].
Carl Linnaeus contributed to the universal acceptance of this principle, constructing species names in his work Species Plantarum in 1753. The scientific names of species, commonly written in Latin, are derived from Greek, Latin, or are formed from Latinized words of other languages. According to the rules of the International Code of Zoological Nomenclature, the correct full name for the domestic bovine species is Bos taurus Linnaeus, 1758, or Bos taurus L., 1758 [Cichocki et al. 2015Cichocki, W., Ważna, A., Cichocki, J., Rajska, E., Jasiński, A., Bogdanowicz, W. (2015). Polskie nazewnictwo ssaków świata [Polish names of the mammals of the world]. Muzeum Inst. Zool. PAN, Warszawa [in Polish]. Google Scholar]. In Poland, the first scientific work in which the species name Bos taurus was given in the book of priest Krzysztof Kluk published in 1779 in Warsaw [Kluk 1779Kluk, K. (1779). Zwierząt domowych i dzikich, osobliwie kraiowych, historyi naturalney początki i gospodarstwo. Tom 1. O zwierzętach ssących [Domestic and wild, particularly native animals: introduction to natural history and farming. Volume 1. On the suckling animals]. Drukarnia J. K. Mości i Rzeczypospolitej u Księży Pijarów, Warszawa [in Polish]. Google Scholar]. Currently, six distinct species of animals are classified for the genus Bos: domestic cattle (Bos taurus Linnaeus, 1758), yak (Bos grunniens Linnaeus, 1766), gayal (Bos frontalis Lambert, 1804), gaur (Bos gaurus Hamilton-Smith, 1827), and banteng (Bos javanicus D`Alton, 1823) with extinct aurochs (Bos primigenius Bojanus, 1827) also included. The term ‘bovine animals’ in a narrow sense refers to representatives of two subspecies of domestic cattle (i.e. the bovine species): Bos taurus Linnaeus, 1758, and Bos indicus Linnaeus, 1758, the latter living in the hot climate zone.
According to archaeological and genetic evidence, the aurochs (Bos primigenius Bojanus, 1827), as the wild progenitor of cattle, was probably domesticated independently in the two, and some believe that even in the three, ancient civilizations during the Neolithic period, i.e. 8–10 thousand years ago. The animal belongs to the most important domesticated animal species in human history. Present-day livestock animals provide a whole range of evidence for the reason why they were domesticated: they produce milk and meat. Additionally hides and skins, hair, bones and other byproducts of slaughter are obtained, and in many parts of the world the animals are still used for transport and draught [Scheu et al. 2015Scheu, A., Powell, A., Bollongino, R., Vigne, J.D., Tresset, A., Çakırlar, C., Benecke, N., Burger J. (2015). The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet., 16–54. https://doi.org/10.1186/s12863-015-0203-2].
Since the domestication of cattle in the Neolithic, the species has also been part of our cultural heritage. Rebuilding its history is an active field of scientific research and contributes to the understanding of the human past. Today, archaeological information is supplemented by genetic data from DNA evidence from maternal, paternal, or autosomal inheritance. New technologies, in particular mitochondrial DNA analysis, provide an alternative source of knowledge and enable the time of domestication to be determined making use of the family tree of modern bovine breeds. The latest genetic data suggests that the mother lines of domestic cattle originate from the Fertile Crescent region, and the cattle of zebu come from the Indus Valley.
Then, together with the mass movement of people there were migrations of cattle, which led to their dispersal in Asia, Africa, Europe and the New World. Different environmental conditions and selection processes led to changes in their appearance and performance, with a variety of production types and breeds. Since the second half of the 20th century, the world's breeding efforts have resulted in an international exchange of bull semen, which has changed the global patterns of genetic diversity of cattle again.
The basic taxonomic unit within the classification system created by Linnaeus is breed. The term razza (race, breed) was used for the first time in Italian by Federico Grisone [1550]Grisone, F. (1550). Gli ordini di cavalcare. Napoli: Giovani Paolo Suganappo. Google Scholar in a work on horse-riding rules. The word race is derived from the Latin radix meaning ‘root’ [Kosowska and Zwolińska-Bartczak 1999Kosowska, B., Zwolińska-Bartczak, I. (1999). Zarys historii zootechniki [Outline of the history of animal husbandry]. Wydaw. AR Wrocław [in Polish]. Google Scholar].
In modern terms, the phrase ‘a bovine breed’ denotes an animal population of the same species which, under the same or similar environmental conditions, has similar inherited characteristics which distinguish it from other breeds of the same species. Moreover, it is sufficiently numerous to be reproduced within the same breed without inbreeding. Briggs and Briggs [1980]Briggs, H.M., Briggs, D.M. (1980). Modern Breeds of Livestock. Fourth Edition. Macmillan Publishing Co. Google Scholar, the authors of Modern Livestock Breeds, defined the breed as “a group of animals that, as a result of breeding and selection, have certain distinguishable characteristics”.
During the last two centuries several kinds of classification have been developed in order to identify cattle breeds. Several criteria have been used, such as coat color, horn size, cranial types, geography, (presumed) origin, and production purpose. In opinion of Felius at al. [2011], “efforts to classify cattle nearly always resulted in a simplification that only described a part of a complex reality. This not only makes such classification largely arbitrary, but also diminishes its usefulness”.
It is estimated that there are currently 1019 local breeds of cattle worldwide [FAO 2015FAO (2015). The Second Report on the State of the World's Animal Genetic Resources for Food and Agriculture, edited by BD, Scherf and D, Pilling. FAO Commission on Genetic Resources for Food and Agriculture Assessments, Rome, http://www.fao.org/3/a-i4787e.pdf. Google Scholar]. Among them, 205 transboundary breeds were used. In 2014 the highest number of bovine breeds were in Europe and the Caucasus (369), Asia (241) and Africa (176). According to Litwińczuk [2011]Litwińczuk, Z. (2011). Ochrona zasobów genetycznych zwierząt gospodarskich [Protection of genetic resources of farm animals]. PWRiL Warszawa [in Polish]. Google Scholar, in 2010 there were 178 local bovine breeds in 24 EU countries, of which 108 (68%) had a population of less than 7,500 cows.
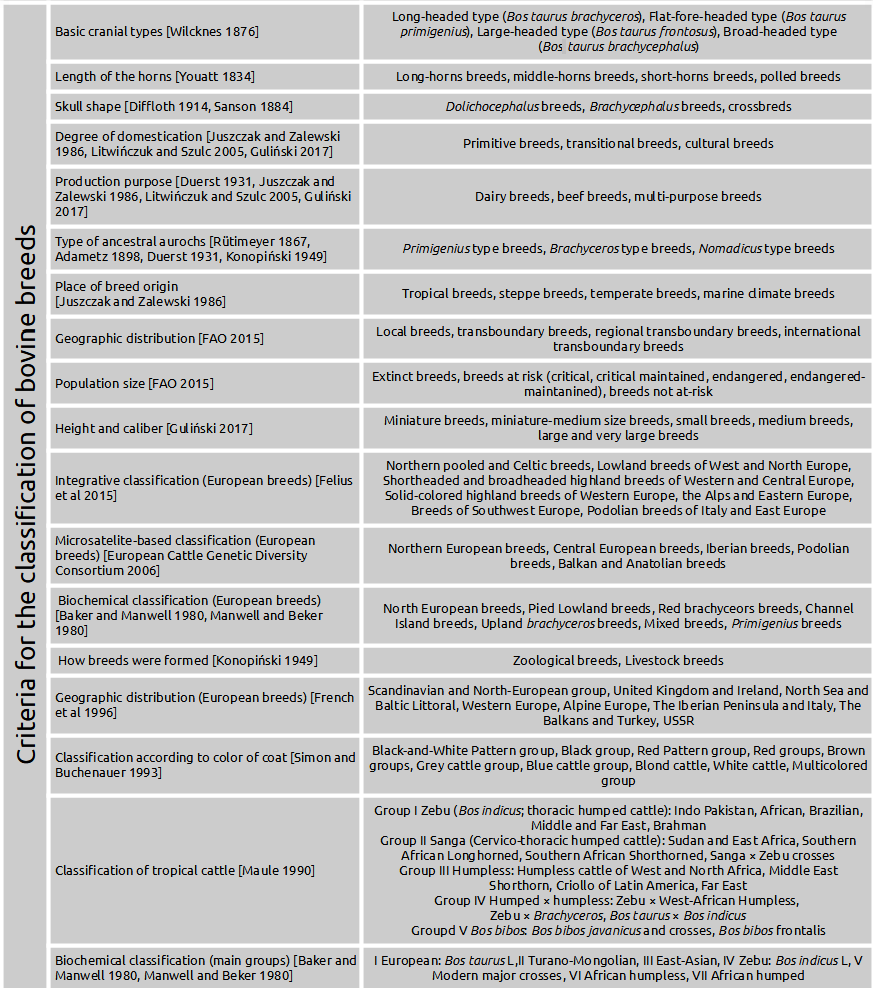
Specialist criteria for the most commonly used classification in modern literature and examples of bovine breeds are presented in Figures 1 and 2. These include basic cranial type, length of the horns, skull shape, color coat, degree of domestication, production purpose, type of ancestral aurochs, geographic distribution, place of breed origin, population size, growth performance, integrative – microsatellite based – biochemical – classifications.
|
Fig. 2. Selected bovine breeds in the world and some criteria for their classification |
At the end of the 19th century, Wilcknes [1876] developed one of the most important criteria for classifying cattle. This author classified cattle into four groups depending on basic cranial types: brachycephalus, primigenius, brachyceros and frontosus. His survey covered only Central-European breeds, a few Dutch and German lowland breeds, the Galloway, Ayrshire, and Shorthorn. He introduced the term brachycephalus after the Bos taurus. Brachyephalus are a cranial type excavated in Italy and dating back to the Roman period. Several scientists modified this classification. Dürst [1899]Dürst, J.U. (1899). Die Rinder von Babylonien, Assyrien und Ägypten und ihr Zusammanhang mit den Rindern der Alten Welt. Georg Reimer, Berlin. https://doi.org/10.5962/bhl.title.86398 also added the long-horned Bos taurus macroceros type to the classification of Wilckens [1876]Wilckens, M. (1876). Die Rinderrassen Mittel-Europas. Wilhelm Braumüller: Wien, Austria. Google Scholar.
A classification of cattle breeds based on the length of horns was presented in paper of Youatt [1834]Youatt, W. (1834). Cattle, Their Breeds, Management and Diseases. Baldwin and Cradock: London, UK. https://doi.org/10.5962/bhl.title.50131. This author distinguished four groups of cattle breeds as follows: the short-, the middle, the long-horns, and polled breeds. It should be emphasized that due to the emergence of genetic technology (CRISPR – Clustered Regularly Interspaced Short Palindromic Repeats) related to the emergence of polled versions of cattle breeds in modern cattle breeding, the classification of breeds based on the use of horns seems to be debatable. Identification of the genetic cause of hornlessness in cattle has been the subject of intensive genetic and genomic research, culminating in the nomination of two different candidate neomutations on cattle chromosome 1 [Carlson et al. 2016Carlson, D.F., Lancto, C.A., Zang, B., Kim, E.S., Walton, M., Oldeschulte, D., Seabury, Ch., Sonstegard, T.S., Fahrenkrug, S. (2016). Production of hornless dairy cattle from genome-edited cell lines. Nat. Biotechnol., 34 (5), 479–481. https://doi.org/10.1038/nbt.3560]. In cattle, the horns allele is genetically recessive to that polled. It is worth noting that in the cattle population, horns trait is far more common in dairy breeds than in beef breeds.
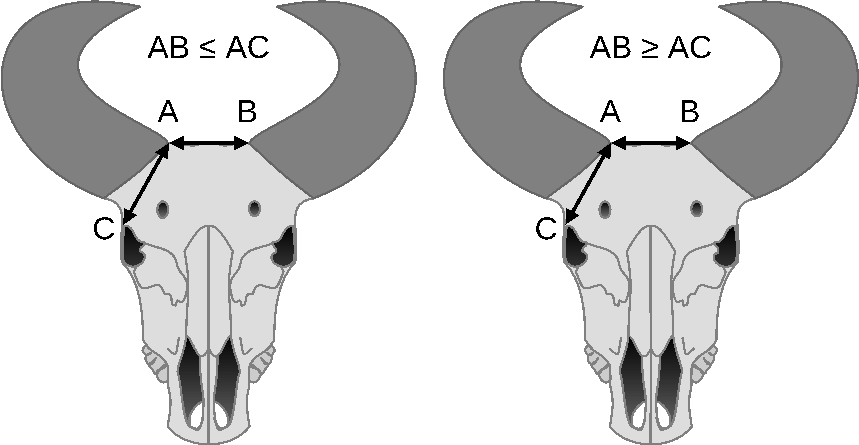
Classification of cattle breeds based on the skull shape supplemented with the assessment of the forms of the poll and horn implant and the length and form of the horns were proposed by Sanson [1884]Sanson, A. (1884). Traité De Zootechnie Tôme 4: Bovidés Taurins Et Bubalins. 3rd ed.; Agricole de las Maison Rustique: Paris, France. Google Scholar and Diffloth [1914]Diffloth, P. (1914). Races Bovines. JB Bailliéres & Fils: Paris, France, 43–44 [in French]. Google Scholar c. This proposal referred to the two basic shapes of the skull that occur in humans, i.e. long-headed people (dolichocephalus) were accompanied by long-skulled cattle and short headed (brachycephaus) people by short-skulled cattle (Fig. 3). In cattle, extreme brachycephaly was described in the Niata extinct breeds of cattle from South America. In the Niata, brachycephaly was considered breed defining even though heavily shortened skulls have been observed in several cattle breeds as a form of malformation [Veitschegger et al. 2018Veitschegger, K., Laura, A.B., Wilson, L.A.B., Nussberger, B., Camenisch, G., Keller, L.F., Wroe, S., Sánchez-Villagra, M.R. (2018). Resurrecting Darwin's Niata – anatomical, biomechanical, genetic, and morphometric studies of morphological novelty in cattle. Sci. Rep., 8, 9129. https://doi.org/10.1038/s41598-018-27384-3].
|
Fig. 3. Classification criteria of bovine breeds into dolichocephalus (AB ≤ AC) and brachycephalus (AB ≥ AC) (own work based on Sanson [1884]Sanson, A. (1884). Traité De Zootechnie Tôme 4: Bovidés Taurins Et Bubalins. 3rd ed.; Agricole de las Maison Rustique: Paris, France. Google Scholar, Diffloth [1914]Diffloth, P. (1914). Races Bovines. JB Bailliéres \& Fils: Paris, France, 43–44 [in French]. Google Scholar) |
The coat color and its pattern seem to be the most obvious breed feature. In principle, within a species, two basic types are distinguished: single-color coats and multicolor coats. The most common single color coats include: white, black, red, yellow, gray, cervine, cream, and brown. Multicolor coats are spotted (black-white, red-white, yellow-white), roan and white-backed. In cattle breeds the coat color is assessed as relevant in many cases. Single-color and multicolor coats are reflected in the names of several bovine breeds (Belgian Blue, Polish Red, Polish Black and White etc.) [Simon and Buchenauer 1993Simon, D.L., Buchenauer, D. (1993). Genetic Diversity of European Livestock Breeds. Wageningen Pres, Wageningen, The Netherlands. Google Scholar].
The first cattle classification systems based on the color of the coat were developed at the turn of the 19th and 20th centuries. In opinion many authors type of coat color was dependent on a particular ethnic group [McKenny Hughes 1896McKenny Hughes, T. (1896). On the more important breeds of cattle which have been recognised in the British Isles in successive periods and their relation to other archaeological and historical discoveries. Archeol., 55, 125–158. https://doi.org/10.1017/S0261340900014296, Kaltenegger 1904Kaltenegger, F. (1904). Prähistorische Und Frühgeschichtlige Periode, I Gruppierung. Wilhelm Frick: Vienna, Austria. Google Scholar, Wilson 1909Wilson, J. (1909). The Evolution of British Cattle and the Fashioning of Breeds. Vinton & Company: Dublin, Ireland. https://doi.org/10.5962/bhl.title.23863]. In the light of this views Roman cattle were to the white, Anglo-Saxon mostly red, Celtic black, and Scandinavian brown-grey. Friesian cows, on the other hand, are known for their black and white coats.
According to the modern knowledge of coat colors, black and red are considered to be the basic colors. Both are associated with one gene – Extension. It is responsible for the production of eumelanin (black pigment) or pheomelanin (red pigment). There are as many as three alleles at the same locus conditioning the two colors.
ED – the dominant allele over the other two, it makes the configuration: EDED, EDe, EDE+, for which the animal is all black. This is the color of such breeds as: Chinese Black Cows or Angus. It is also one of the many other coat colors of such breeds as Brangus, Highland, Simmental, Salers, or Holstein-Frisian.
e – recessive allele, which in the configuration of ee or E+e determines a lack of the black pigment, so the animals are all red. Such a color can be found in Hereford, Red Angus, and Polish Red breeds.
E+ – this version of the gene called wild type has a neutral effect in configuration with the ED or e allele. Both black and red pigments are produced in the E+E+ system. This can be observed in Swiss Brown, Steppe, and Jersey cattle, and in primitive breeds of Europe and the Middle East. The coat color in cattle is inherited according to the Pisum pattern based on total domination. Black coat is conditioned by allele A, red by a, single-color by B, and spotted coat by b [Gołębiewski and Brzozowski 2007Gołębiewski, M., Brzozowski, P. (2007). Genetyka umaszczenia bydła [Color coat of cattle genetics]. Pr. Mat. Zootech., 64, 7–21 [in Polish]. Google Scholar].
The most important characteristics determining the classification of bovine breeds are production purpose. Duerst [1931]Duerst, J.U. (1931). Grundlagen der Rinderzucht. Springer Verlag, Berlin. https://doi.org/10.1007/978-3-662-28664-7 introduced the concept of respiratory and digestive type in cattle, providing the basis for the classification of breeds into dairy and meat. By measuring the angle of the last rib, he obtained in practice a simple way to divide the types of cattle into respiratory (dairy), combined respiratory and digestive, combined digestive and digestive (meat). Currently, in developed countries, cattle are reared for three basic production purposes: either dairy or meat or both. Dairy breeds of bovine animals include those used for the production of large quantities of milk intended for consumption or processing by dairies. It should be stressed that today's highly productive dairy breeds in the world belong exclusively to the Bos taurus, L. subspecies.
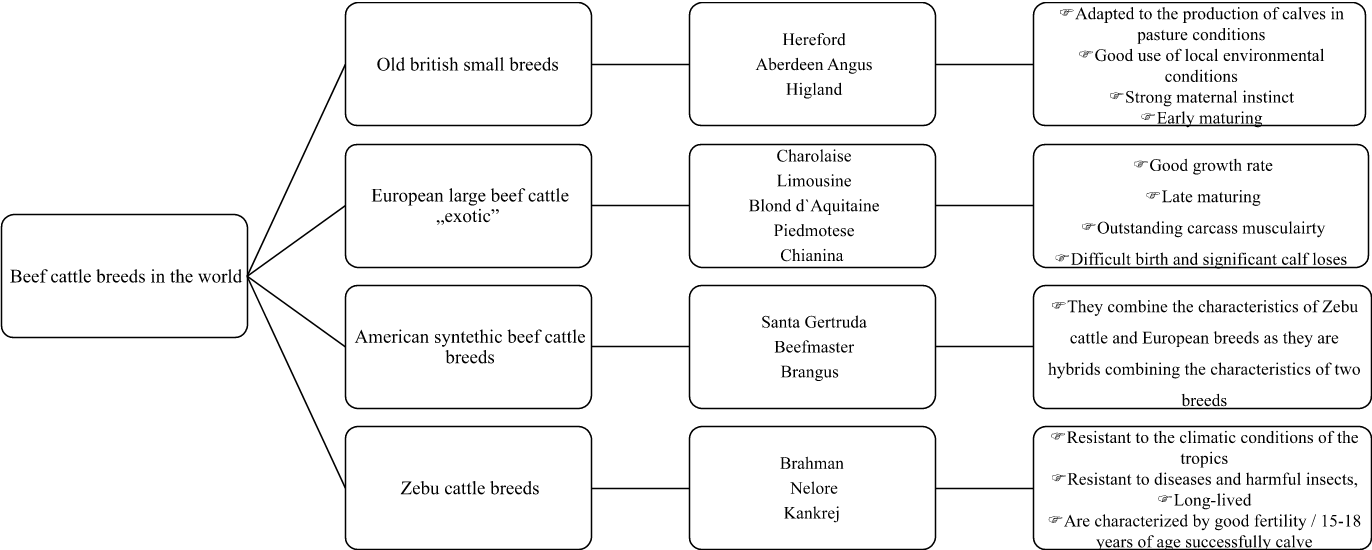
Until the beginning of the 20th century, farmers bred dual-purpose cattle; the same herd was used for both milk and meat production. Today, in developed countries with capitalist economy, cattle are bred to produce either milk or beef. The efficiency of the business and the quality of food, enforced by the market, has also led to specialization in both milk and beef processing sectors. There are only a few breeds of dairy cattle in the world. These breeds are Holstein [Holstein Association USA 2020Holstein Association USA (2020). http://www.holsteinusa.com.. Google Scholar], in Poland, Polish Holstein-Frisian, Brown Swiss, Guernsey, Ayrshire, Jersey, Shorthorn Dairy, and Montbeliarde. Bovine breeds raised for beef production are also called beef cattle. Animals belonging to this production type have a rectangular body shape and a strong, deep chest. The ribs are narrowly spaced at the right angle in relation to the spine. They also have thick spongy skin and are well muscled in particular in the loin and rump. Animals of this production type are of a calm, phlegmatic temperament and reduced metabolism. Today, nearly 250 beef cattle breeds are used in meat production worldwide. The most important in terms of population size and economy are Hereford, Charolaise, Aberdeen Angus, Limousine and Meat Shorthorn. The group of beef breeds is the largest within the species. Figures 4 and 5 lists groups of beef breeds classified according to the place of their creation and height and caliber.
|
Fig. 4. Classification beef breeds according to the place of origin |
|
Fig. 5. Classification beef breeds according to the height and caliber |
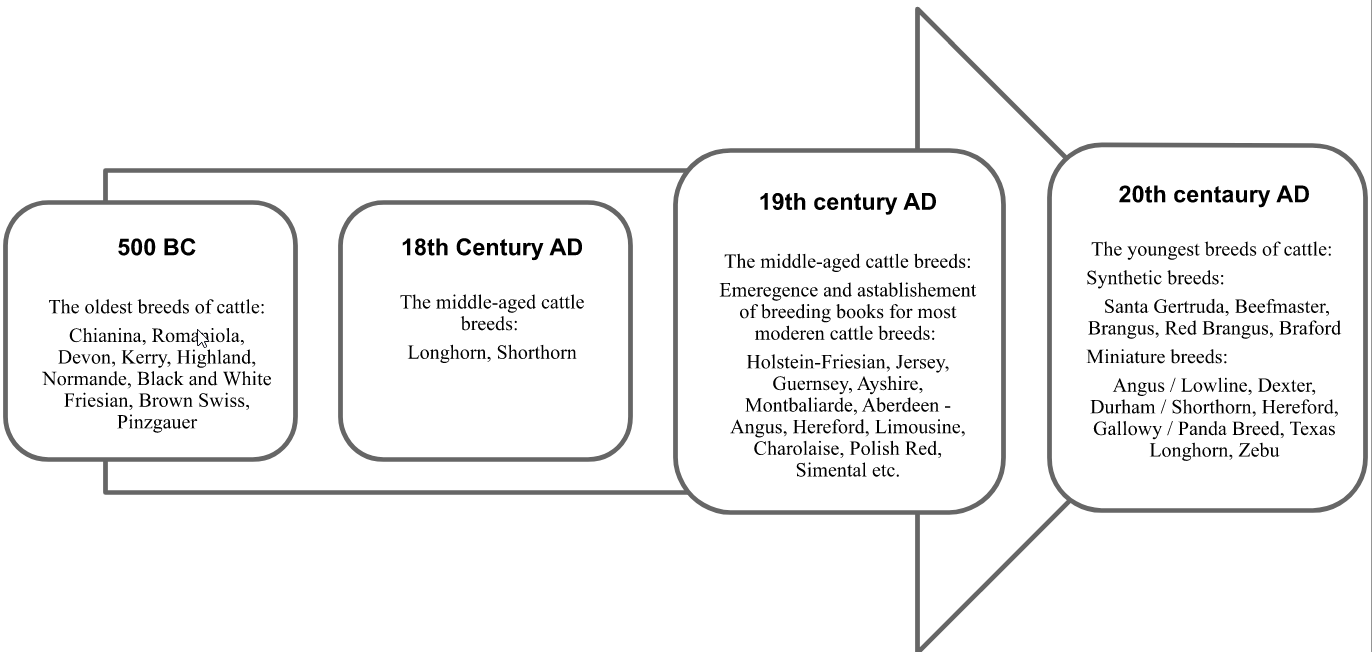
Domestic cattle have been reared by humans for thousands of years. Selective breeding of these animal species was used to modify its characteristics. To achieve that, different types of scientific knowledge was applied, but selection was and still remains the basic technique used in breeding. It involves mating animals deliberately selected in terms of expected characteristics. Cattle farmers have used selection for thousands of years. This has led to the development of hundreds of breeds of cattle, with some of them already existing in ancient times. The Italian Chianina and Romaniola breeds, whose history dates back to the Roman Empire, are widely regarded as some of the oldest (Fig. 6). Black and white Frisian cows also have a long history. The beginning of breeding black and white Holstein-Frisian cattle dates back to around 100 BC. The Frisians were a Germanic tribe living on the North Sea coast, currently in the northern Netherlands and Germany. The term originates from frees/fries, an Old Germanic word denoting ‘edge’ or ‘shore’. Thus, literally, the word Frisian means the inhabitant of the coast. The Frisians were known as successful cattle breeders. Their cattle were a result of crossbreeding of primitive breeds with characteristic black and white coats. The Frisians bred black and white cows for 2,000 years in purity, without any crossbreeding. The development of cattle farms in the region was facilitated by polder areas, very abundant in grassland. The breeding, improvement and maintenance of cattle has resulted in great success [Guliński 2017Guliński, P. (2017). Bydło domowe hodowla i użytkowanie [Domestic Cattle Breeding and Management]. Wyd. Nauk. PWN, Warszawa [in Polish]. Google Scholar, Litwińczuk and Guliński 2000Litwińczuk, Z., Guliński, P. (2000). Bydło holsztyńsko-fryzyjskie i jego wykorzystanie w doskonaleniu czarno-białego bydła mlecznego w Polsce i na świecie [Holstein-Friesian cattle and their use in the improvement of Black-and-White dairy cattle in Poland and world-wide]. Post. Nauk Rol., 2, 71–87 [in Polish with an abstract in English]. Google Scholar].
|
Fig. 6. Selected bovine breeds according to time of their emergence |
Cattle breeds can be also classified based on types of aurochs they descend from. Swiss researcher Rütimeyer [1867]Rütimeyer, L. (1867). Versuch einer natürlichen Geschichte des Rindes, in seinen Beziehungen zu den Wiederkauern im Allgemeinen. Druck von Zürcher und Furrer, Zürich, Switherland. Google Scholar identified two aurochs species: Bos primigenius (Bojanus, 1827) and an early form of Indian aurochs called Bos nomadicus [Falconer 1868Falconer, H. (1868). Paleontological Memoirs and Notes of the Late Hugh Falconer; Vol. II. Robert Hardwicke, London. Google Scholar]. He also held that shorthorned cattle were the oldest and most widespread form of domestic cattle (Bos taurus) of Neolithic Europe. Contrary to that, Adametz [1898]Adametz, L. (1898). Studien über Bos (brachyceros) europaeus, die wilde Stamform der Brachycerosrassen der europäischen Hausrindes. J. Landwirt., 48, 17–31. Google Scholar considered Bos brachyceros to have been a genuine European wild form. The breeds belonging to the primigenius type share a strong build and high growth, as well as the presence of horns. Their skull structure is characterized by a long face, a flat forehead, eye sockets located in the forehead plane, deep and narrow temporal pits, wide and low occiput. The breed of cattle of the brachyceros type is characterized by a smaller height and caliber than that of primigenius cattle and less developed horns [Duerst 1931Duerst, J.U. (1931). Grundlagen der Rinderzucht. Springer Verlag, Berlin. https://doi.org/10.1007/978-3-662-28664-7]. The facial part of the skull is shorter, the forehead is cup-shaped, the arches of the eye sockets protrude above the forehead plane, the temporal pits are shallow and wide, and the poll is narrow and high. The structure of the nomadicus cattle shows the characteristics of zebu cattle.
Juszczak and Zalewski [1986]Juszczak, J., Zalewski, W. (1986). Hodowla bydła [Cattle breeding]. PWRiL Warszawa [in Polish]. Google Scholar assume that the origin of cattle breeds goes back to three different types of aurochs. Therefore, they distinguish three types of cattle breeds: primigenius, brachyceros, and nomadicus. Breeds of cattle of the primigenius type combine strong structure and high growth performance, as well as the presence of horns. In the construction of their skull, a characteristic feature is a long facial part, a flat forehead, eye socket arches located in the plane of the forehead, deep and narrow temporal fossa, with occiput wide and low. Bovine breeds of the brachyceros type are of a smaller growth performance and size than primigenius cattle and with less developed horns. The facial part of the skull is shorter, the forehead is recessed, the arches of the eye sockets protrude above the plane of the forehead, the temporal fossa is shallow and wide, and the occiput narrow and high. The nomadicus breeds in their construction are similar to zebu cattle.
One of the most important criteria for the classification of cattle breeds is their geographic distribution. Taking into account this criterion, contemporary cattle breeds can be classified into the following groups: local breeds (breeds that occur only in one country), transboundary breeds (breeds that occur in more than one country), regional transboundary breeds (transboundary breeds that occur only in one of the seven SoW-AnGR regions) and international transboundary breeds (transboundary breeds that occur in more than one SoW-AnGR region) [FAO 2015FAO (2015). The Second Report on the State of the World's Animal Genetic Resources for Food and Agriculture, edited by BD, Scherf and D, Pilling. FAO Commission on Genetic Resources for Food and Agriculture Assessments, Rome, http://www.fao.org/3/a-i4787e.pdf. Google Scholar]. Seven regions were defined for the SoW-AnGR: Africa, Asia, Europe and the Caucasus, Latin America and the Caribbean, the Near and Middle East, North America and the Southwest Pacific.
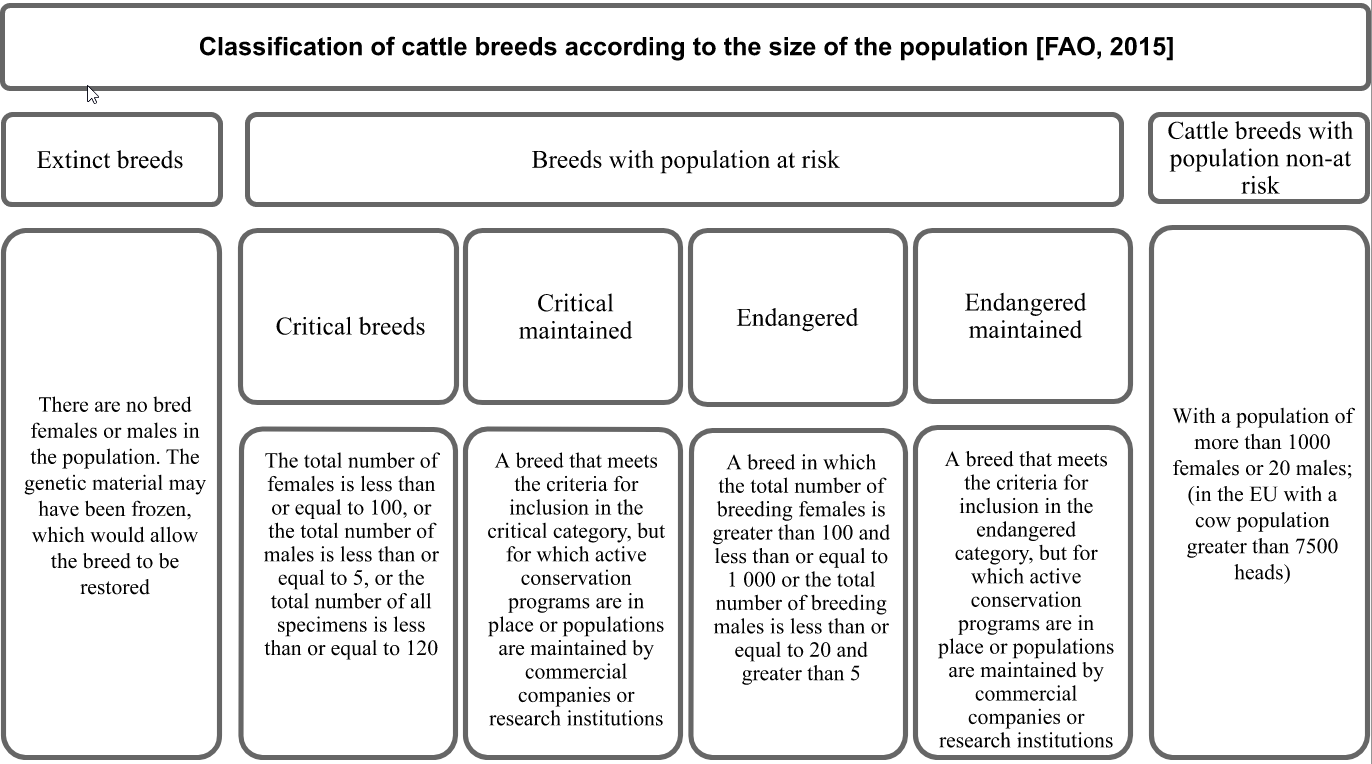
Another important criterion that is often used to classify today's bovine breeds is the size of their population. The actual FAO classification [FAO 2015FAO (2015). The Second Report on the State of the World's Animal Genetic Resources for Food and Agriculture, edited by BD, Scherf and D, Pilling. FAO Commission on Genetic Resources for Food and Agriculture Assessments, Rome, http://www.fao.org/3/a-i4787e.pdf. Google Scholar] divides the cattle breed into two basic groups: at risk and not at risk. Within the group of at risk, the following groups of breeds are distinguished: extinct, critical, critical-maintained, endangered and endangered-maintained. The rules for the division of bovine breeds according to the this criterion are set out in Fig. 7.
|
Fig. 7. Classification bovine breeds according to the size of population [FAO 2015]FAO (2015). The Second Report on the State of the World's Animal Genetic Resources for Food and Agriculture, edited by BD, Scherf and D, Pilling. FAO Commission on Genetic Resources for Food and Agriculture Assessments, Rome, http://www.fao.org/3/a-i4787e.pdf. Google Scholar |
Genetic development of cattle breeding was closely linked to technological progress in agriculture. Becoming popular since the First World War, tractors have gradually replaced horses and cattle in agriculture in developed countries. Consequently, in 1950 there was a change in the breeding objective, from cattle producing meat, milk, but also used for draught and transport purposes, to animals providing milk and meat only. Old triple-purpose (meat/milk/draft) native cattle types that had been raised for centuries began to decline, and in their place, more productive double- and single-purpose cattle were bred. The introduction of milking machines in the 1960s intensified the development of specialized dairy breeds, from which the correct construction of the udder, suitable for mechanical milking, was expected. The use of modern breeding technologies, such as artificial insemination since the 1930s and embryo transfer since the 1970s, has allowed a wide spread of breeding material around the world. In this way, it was possible to use the breeding material of different breeds of cattle on an unprecedented scale, in different countries, and on different continents. However, these changes had an obvious drawback. The focus on the most productive breeds was at the expense of less efficient local cattle breeds. Many of them disappeared or were crossed to such an extent that they were practically extinct even though those breeds had a promising potential with their ability to withstand local, sometimes extreme, environmental conditions. The loss of local breeds can also lead to erosion of genetic resources that are difficult to replace [Hiemstra et al. 2010Hiemstra, S.J., de Haas, Y., Mäki-Tanila, A., Gandini, G. (2010). Local cattle breeds in Europe. Development of policies and strategies for self-sustaining breeds. Wageningen Academic Publishers, The Netherlands. https://doi.org/10.3920/978-90-8686-697-7, Guliński 2017Guliński, P. (2017). Bydło domowe hodowla i użytkowanie [Domestic Cattle Breeding and Management]. Wyd. Nauk. PWN, Warszawa [in Polish]. Google Scholar]. Currently, in Europe, the most famous local breeds of cattle include: in France – Ferrandaise and Villard de Lans, Abondance, Tarentaise, Armorican; in Italy – Reggiana and Modenese; in the Netherlands – Groningen White-Headed and Meuse Rhine Yssel; in Finland – Western Finncattle; in Spain – Alistana Sanabresa, Pajuna, Limia; in Poland – Polish Red, Polish Black and White, Polish Red and White; in Ireland – Kerry Cattle and Irish Moiled.
The growth performance of animals in modern cattle farming is most often determined by the height (cm) at the highest point of the back, i.e. withers or rump, during full somatic development. In cattle, this is considered to be 3–5 years of age. Table 1 provides information on the diversity of modern bovine breeds’ growth performance.
Table 1. Bovine breeds according to their growth performance |
|
Breeds group |
Height at rump of a female at the age of 3 years, cm |
Miniature cattle breeds |
0–105 |
Medium–sized miniature cattle breeds |
105–120 |
Breeds of small cattle |
120–130 |
Medium breeds of cattle |
130–145 |
Breeds of large and very large cattle |
145–180 |
According to these data, present-day domestic cattle include very diverse breeds in terms of growth performance and size, ranging from miniature breeds, reaching up to 105 cm in height at the rump, to very large animals, reaching the height at the rump of above 145 cm at the age of full somatic development. According to archaeological data from Ostrów Lednicki, the average height at the withers of cattle in the Middle Ages in Poland during the time of Mieszko I (10th century AD) was only 104 cm [Makowiecki 2001Makowiecki, D. (2001). Hodowla oraz użytkowanie zwierząt na Ostrowie Lednickim w średniowieczu. Studium archeozoologiczne [Breeding and use of animals in Ostrów Lednicki in the Middle Ages. Archaeozoological study]. Bibl. Stud. Ledn., Poznań, VI [in Polish]. Google Scholar]. It should also be noted that the selection programs for economically important bovine breeds in the 20th and 21st centuries have aimed to increase their size and growth performance. This applies to both dairy cattle (the Holstein breed in the USA and Holstein-Frisian in Poland) and meat bovines (continental and American breeds of synthetic cattle). The reason for this is that growth performance positively affects a number of other economically important characteristics.
Konopiński [1949]Konopiński, T. (1949). Hodowla bydła [Cattle breeding]. Inst. Wydaw. Ruchu Lud. ,,Polska'', Poznań [in Polish]. Google Scholar used classification criteria according to how breeds were formed. The first group was constituted by breeds whose formation was mostly affected by the environment and living conditions, in the second group he included those developed by human activity, with the production purpose playing a key role. He described the first group of breeds as ‘zoological’. The second group are man-made breeds, or so-called livestock breeds, a population of homozygous animals in terms of certain characteristics, usually associated with the purpose of cattle farming. Zoological breeds are of a very balanced genotype (long development time of the breed) and more developed abilities of constant transmission of traits. Breeding breeds show small, because only in terms of a few features, the alignment of genotypes, and a less outstanding ability to constantly transfer features to offspring.
According to Felius [1985]Felius, M. (1985). Genus Bos: cattle breeds in the world. MSDAGVET Rahway, USA. Google Scholar the genus Bos can be divided into 16 groups, with major ones briefly described by the author (with origin, color, size and production purpose). The first five are all western European breeds (202 breeds). Two groups of those 16 are reared in southeast Europe, the Caucasus, Asia Minor and the Middle East (41 breeds), two more are Asian breeds (72 breeds), five groups are African breeds (108 breeds), and the last two are Criollo cattle (38 breeds) and new breeds (19 breeds). Each group is divided into two or more subgroups, largely on the basis of relationship or similarity within each subgroup.
In 1995, Felius [1995]Felius, M. (1995). Cattle Breeds: An Encyclopedia. Trafalgar Square Publishing. Google Scholar developed a comprehensive classification of bovine domestic breeds. This classification is based on morphological, geographical and historical data. Of the three criteria for classification, geography was proposed to be the most important. The breeds have been arranged first according to continental origin, which is plausible because cattle from different continents are likely to have developed relatively independently (isolation by distance).
Based on protein polymorphism data (α-lactalbumin, β-lactoglobulin, caseins (αsl, β and x), serum albumin, transferrin, haemoglobin, amylase and carbonic anhydrase) Baker and Manwell [1980]Baker, C.M., Manwell, C. (1980). Chemical classification of cattle. 1. Breed groups. Anim. Blood Groups Biochem. Gen., 11, 127–150. https://doi.org/10.1111/j.1365-2052.1980.tb01503.x and Manwell and Baker [1980]Manwell, C., Baker, C.M. (1980). Chemical classification of cattle. 2. Phylogenetic tree and specific status of the zebu. Anim. Blood Groups Biochem. Gen., 11, 151–162. https://doi.org/10.1111/j.1365-2052.1980.tb01504.x compared the frequency of alleles in 196 different breeds of cattle worldwide. The results of their work made it possible to distinguish 7 groups of cattle in the world. Their names are given in Fig. 1. Within the European cattle, the authors distinguished the following groups: Primigenius, primigenius-brachyceros Mixed, Upland brachyceros, Channel Island brachyceros, European Red brachyceros, Pied Lowland and North-Scandinavian [Baker and Manwell 1980Baker, C.M., Manwell, C. (1980). Chemical classification of cattle. 1. Breed groups. Anim. Blood Groups Biochem. Gen., 11, 127–150. https://doi.org/10.1111/j.1365-2052.1980.tb01503.x, Manwell and Baker 1980Manwell, C., Baker, C.M. (1980). Chemical classification of cattle. 2. Phylogenetic tree and specific status of the zebu. Anim. Blood Groups Biochem. Gen., 11, 151–162. https://doi.org/10.1111/j.1365-2052.1980.tb01504.x].
The development of genetic technologies related to DNA analysis observed in the last 20 years has obviously also influenced the analysis of the origin of different breeds of cattle, which are compared using several types of genetic markers at the DNA level. Various variants of mitochondrial DNA in the form of microsatellites and single nucleotide polymorphisms (ASNPs) are selected as genetic markers. The analysis of the structure of these markers makes it possible in practice to determine the similarity or lack thereof between the compared groups of cattle breeds [Groeneveld et al. 2010Groeneveld, L.F., Lenstra, J.A., Eding, H., Toro, M.A., Scherf, B., Pilling, D., Negrini, R., Finlay, E. K., Jianlin, H., Groeneveld, E., Weigend, S. (2010). Genetic diversity in farm animals: A review. Anim. Genet., 41, 6–31. https://doi.org/10.1111/j.1365-2052.2010.02038.x]. From the research on the use of genetic markers to analyze the origin of cattle, research funded by the European Commission and carried out at the beginning of the 21st century in laboratories of several European countries deserve emphasis. These studies analyzed all major and selected local breeds of cattle kept in Europe. In total, 69 breeds of cattle were analyzed. The results of this extensive work, coupled with the development of phylogenetic networks, revealed the common ancestry of four groups of European cattle breeds. These were the following breed groups: Northern, Central, Iberian and Podolian cattle. There were also significant differences in the structure of the analyzed markers for primitive cattle breeds kept in the Balkans and Anatolia [Felius et al. 2011Felius, M., Koolmees, P.A., Theunissen, B., Lenstra, J.A. (2011). On the Breeds of Cattle-Historic and Current Classifications. Diversity. European Cattle Genetic Diversity Consortium, 3(4), 660–692. https://doi.org/10.3390/d3040660].
The data presented in the paper on the origin of cattle and the criteria for classifying breeds belonging to this species of animals are the basis for the following summaries and reflections. Nowadays, the importance of knowledge in this field should be considered on two levels: theoretical – cognitive and practical – application. Of course, knowledge about the origin of cattle, the processes of its domestication and the formation of cattle breeds has a cognitive value that is important in scientific activity and in the transfer of knowledge at its various levels. It seems that the use of knowledge about the systems and criteria for the classification of cattle breeds is of particular importance. The following justification should be indicated for a positive assessment of the issues undertaken in the work. Classifying cattle breeds in practical terms contributes to solving a number of problems occurring in cattle breeding, which include e.g. proper selection of the breed to the environmental and economic conditions or activities related to the protection of genetic resources. It should be assumed that, due to the not always high level of knowledge of breeders, some classification criteria, such as the purpose type, growth rate, and population size, play a fundamental role in the selection of specific breeds for the production of a specific type of food. Knowledge of the criteria for the classification of cattle breeds is also one of the basic issues in teaching the subject of cattle breeding at a higher level. Achieving learning outcomes in this field is an important criterion for achieving the qualifications and competences of agricultural students.
This study was financed by the funds of the Ministry of Science and Higher Education of Poland (statutory research fund of the Siedlce University of Natural Sciences and Humanities No 50/20/B).
Received: 16 Jul 2021
Accepted: 28 Sep 2021
Published online: 27 Nov 2021
Accesses: 2730
Guliński, P., (2021). Cattle breeds – contemporary views on their origin and criteria for classification: a review. Acta Sci. Pol. Zootechnica, 20(2), 3–18. DOI: 10.21005/asp.2021.20.2.01.